
Abstract
The pathway causing CD4 T-cell death in HIV-infected hosts remains poorly understood although apoptosis has been proposed as a key mechanism. We now show that caspase-3-mediated apoptosis accounts for the death of only a small fraction of CD4 T cells corresponding to those that are both activated and productively infected. The remaining over 95% of quiescent lymphoid CD4 T cells die by caspase-1-mediated pyroptosis triggered by abortive viral infection. Pyroptosis corresponds to an intensely inflammatory form of programmed cell death in which cytoplasmic contents and pro-inflammatory cytokines, including IL-1β, are released. This death pathway thus links the two signature events in HIV infection—CD4 T-cell depletion and chronic inflammation—and creates a pathogenic vicious cycle in which dying CD4 T cells release inflammatory signals that attract more cells to die. This cycle can be broken by caspase 1 inhibitors shown to be safe in humans, raising the possibility of a new class of ‘anti-AIDS’ therapeutics targeting the host rather than the virus.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others
References
Thomas, C. Roadblocks in HIV research: five questions. Nature Med. 15, 855–859 (2009)
Muro-Cacho, C. A., Pantaleo, G. & Fauci, A. S. Analysis of apoptosis in lymph nodes of HIV-infected persons. Intensity of apoptosis correlates with the general state of activation of the lymphoid tissue and not with stage of disease or viral burden. J. Immunol. 154, 5555–5566 (1995)
Finkel, T. H. et al. Apoptosis occurs predominantly in bystander cells and not in productively infected cells of HIV- and SIV-infected lymph nodes. Nature Med. 1, 129–134 (1995)
Huang, M. B., James, C. O., Powell, M. D. & Bond, V. C. Apoptotic peptides derived from HIV-1 Nef induce lymphocyte depletion in mice. Ethnic. Dis. 18, S2–30–37 (2008)
Røsok, B. I. et al. Correlates of apoptosis of CD4+ and CD8+ T cells in tonsillar tissue in HIV type 1 infection. AIDS Res. Hum. Retroviruses 14, 1635–1643 (1998)
Gougeon, M. L. et al. Programmed cell death in peripheral lymphocytes from HIV-infected persons: increased susceptibility to apoptosis of CD4 and CD8 T cells correlates with lymphocyte activation and with disease progression. J. Immunol. 156, 3509–3520 (1996)
Jekle, A. et al. In vivo evolution of human immunodeficiency virus type 1 toward increased pathogenicity through CXCR4-mediated killing of uninfected CD4 T cells. J. Virol. 77, 5846–5854 (2003)
Grivel, J. C., Malkevitch, N. & Margolis, L. Human immunodeficiency virus type 1 induces apoptosis in CD4+ but not in CD8+ T cells in ex vivo-infected human lymphoid tissue. J. Virol. 74, 8077–8084 (2000)
Lamkanfi, M. & Dixit, V. M. Manipulation of host cell death pathways during microbial infections. Cell Host Microbe 8, 44–54 (2010)
Cooper, A. et al. HIV-1 causes CD4 cell death through DNA-dependent protein kinase during viral integration. Nature 498, 376–379 (2013)
Doitsh, G. et al. Abortive HIV infection mediates CD4 T cell depletion and inflammation in human lymphoid tissue. Cell 143, 789–801 (2010)
Eckstein, D. A. et al. HIV-1 actively replicates in naive CD4+ T cells residing within human lymphoid tissues. Immunity 15, 671–682 (2001)
Glushakova, S., Baibakov, B., Margolis, L. B. & Zimmerberg, J. Infection of human tonsil histocultures: a model for HIV pathogenesis. Nature Med. 1, 1320–1322 (1995)
Ren, Y. & Savill, J. Apoptosis: the importance of being eaten. Cell Death Differ. 5, 563–568 (1998)
Fink, S. L. & Cookson, B. T. Apoptosis, pyroptosis, and necrosis: mechanistic description of dead and dying eukaryotic cells. Infect. Immun. 73, 1907–1916 (2005)
Levy, D. N., Aldrovandi, G. M., Kutsch, O. & Shaw, G. M. Dynamics of HIV-1 recombination in its natural target cells. Proc. Natl Acad. Sci. USA 101, 4204–4209 (2004)
Bedner, E., Smolewski, P., Amstad, P. & Darzynkiewicz, Z. Activation of caspases measured in situ by binding of fluorochrome-labeled inhibitors of caspases (FLICA): correlation with DNA fragmentation. Exp. Cell Res. 259, 308–313 (2000)
Collman, R. et al. An infectious molecular clone of an unusual macrophage-tropic and highly cytopathic strain of human immunodeficiency virus type 1. J. Virol. 66, 7517–7521 (1992)
Mariathasan, S. et al. Cryopyrin activates the inflammasome in response to toxins and ATP. Nature 440, 228–232 (2006)
Schroder, K. & Tschopp, J. The inflammasomes. Cell 140, 821–832 (2010)
Netea, M. G. et al. Differential requirement for the activation of the inflammasome for processing and release of IL-1β in monocytes and macrophages. Blood 113, 2324–2335 (2009)
Laliberte, R. E., Eggler, J. & Gabel, C. A. ATP treatment of human monocytes promotes caspase-1 maturation and externalization. J. Biol. Chem. 274, 36944–36951 (1999)
Perregaux, D. & Gabel, C. A. Interleukin-1β maturation and release in response to ATP and nigericin. Evidence that potassium depletion mediated by these agents is a necessary and common feature of their activity. J. Biol. Chem. 269, 15195–15203 (1994)
Perregaux, D. et al. IL-1β maturation: evidence that mature cytokine formation can be induced specifically by nigericin. J. Immunol. 149, 1294–1303 (1992)
Moore, J. P., Kitchen, S. G., Pugach, P. & Zack, J. A. The CCR5 and CXCR4 coreceptors–central to understanding the transmission and pathogenesis of human immunodeficiency virus type 1 infection. AIDS Res. Hum. Retroviruses 20, 111–126 (2004)
Schweighardt, B. et al. R5 human immunodeficiency virus type 1 (HIV-1) replicates more efficiently in primary CD4+ T-cell cultures than X4 HIV-1. J. Virol. 78, 9164–9173 (2004)
Grivel, J. C. & Margolis, L. B. CCR5- and CXCR4-tropic HIV-1 are equally cytopathic for their T-cell targets in human lymphoid tissue. Nature Med. 5, 344–346 (1999)
Zhou, Y., Shen, L., Yang, H. C. & Siliciano, R. F. Preferential cytolysis of peripheral memory CD4+ T cells by in vitro X4-tropic human immunodeficiency virus type 1 infection before the completion of reverse transcription. J. Virol. 82, 9154–9163 (2008)
Lanzavecchia, A. & Sallusto, F. Dynamics of T lymphocyte responses: intermediates, effectors, and memory cells. Science 290, 92–97 (2000)
Mackay, C. R. Immunological memory. Adv. Immunol. 53, 217–265 (1993)
Sallusto, F., Lenig, D., Forster, R., Lipp, M. & Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 401, 708–712 (1999)
Bergsbaken, T., Fink, S. L. & Cookson, B. T. Pyroptosis: host cell death and inflammation. Nature Rev. Microbiol. 7, 99–109 (2009)
Decker, T. & Lohmann-Matthes, M. L. A quick and simple method for the quantitation of lactate dehydrogenase release in measurements of cellular cytotoxicity and tumor necrosis factor (TNF) activity. J. Immunol. Methods 115, 61–69 (1988)
Ventura, A. et al. Cre-lox-regulated conditional RNA interference from transgenes. Proc. Natl Acad. Sci. USA 101, 10380–10385 (2004)
Baldauf, H. M. et al. SAMHD1 restricts HIV-1 infection in resting CD4+ T cells. Nature Med. 18, 1682–1687 (2012)
Agosto, L. M. et al. The CXCR4-tropic human immunodeficiency virus envelope promotes more-efficient gene delivery to resting CD4+ T cells than the vesicular stomatitis virus glycoprotein G envelope. J. Virol. 83, 8153–8162 (2009)
Boxer, M. B., Shen, M., Auld, D. S., Wells, J. A. & Thomas, C. J. A small molecule inhibitor of Caspase-1. in Probe Reports from the NIH Molecular Libraries Program (Bethesda Maryland, 2010)
Boxer, M. B. et al. A highly potent and selective caspase-1 inhibitor that utilizes a key 3-cyanopropanoic acid moiety. ChemMedChem 5, 730–738 (2010)
Randle, J. C., Harding, M. W., Ku, G., Schonharting, M. & Kurrle, R. ICE/Caspase-1 inhibitors as novel anti-inflammatory drugs. Expert Opin. Investig. Drugs 10, 1207–1209 (2001)
Stack, J. H. et al. IL-converting enzyme/caspase-1 inhibitor VX-765 blocks the hypersensitive response to an inflammatory stimulus in monocytes from familial cold autoinflammatory syndrome patients. J. Immunol. 175, 2630–2634 (2005)
Maroso, M. et al. Interleukin-1β biosynthesis inhibition reduces acute seizures and drug resistant chronic epileptic activity in mice. Neurotherapeutics 8, 304–315 (2011)
Vezzani, A. et al. ICE/caspase 1 inhibitors and IL-1β receptor antagonists as potential therapeutics in epilepsy. Current Opinion in Investigational Drugs 11, 43–50 (2010)
Février, M., Dorgham, K. & Rebollo, A. CD4+ T cell depletion in human immunodeficiency virus (HIV) infection: role of apoptosis. Viruses 3, 586–612 (2011)
Monroe et al. IFI16 DNA sensor is required for death of lymphoid CD4 T cells abortively infected with HIV. Science http://dx.doi.org/10.1126/science.1243640 (19 December 2013)
Biancotto, A. et al. HIV-1 induced activation of CD4+ T cells creates new targets for HIV-1 infection in human lymphoid tissue ex vivo. Blood 111, 699–704 (2008)
Zeng, M. et al. Cumulative mechanisms of lymphoid tissue fibrosis and T cell depletion in HIV-1 and SIV infections. J. Clin. Invest. 121, 998–1008 (2011)
Deeks, S. G. HIV infection, inflammation, immunosenescence, and aging. Annu. Rev. Med. 62, 141–155 (2011)
Papkalla, A., Munch, J., Otto, C. & Kirchhoff, F. Nef enhances human immunodeficiency virus type 1 infectivity and replication independently of viral coreceptor tropism. J. Virol. 76, 8455–8459 (2002)
Goujon, C. et al. With a little help from a friend: increasing HIV transduction of monocyte-derived dendritic cells with virion-like particles of SIV(MAC). Gene Ther. 13, 991–994 (2006)
Laguette, N. et al. SAMHD1 is the dendritic- and myeloid-cell-specific HIV-1 restriction factor counteracted by Vpx. Nature 474, 654–657 (2011)
Wissing, S., Montano, M., Garcia-Perez, J. L., Moran, J. V. & Greene, W. C. Endogenous APOBEC3B restricts LINE-1 retrotransposition in transformed cells and human embryonic stem cells. J. Biol. Chem. 286, 36427–36437 (2011)
Grivel, J. C. et al. HIV-1 pathogenesis differs in rectosigmoid and tonsillar tissues infected ex vivo with CCR5- and CXCR4-tropic HIV-1. AIDS 21, 1263–1272 (2007)
Wigler, M., Pellicer, A., Silverstein, S. & Axel, R. Biochemical transfer of single-copy eucaryotic genes using total cellular DNA as donor. Cell. 14, 725–731 (1978)
Doranz, B. J. et al. A dual-tropic primary HIV-1 isolate that uses fusin and the beta-chemokine receptors CKR-5, CKR-3, and CKR-2b as fusion cofactors. Cell 85, 1149–1158 (1996)
De Rosa, S. C., Herzenberg, L. A., Herzenberg, L. A. & Roederer, M. 11-color, 13-parameter flow cytometry: identification of human naive T cells by phenotype, function, and T-cell receptor diversity. Nature Med. 7, 245–248 (2001)
Brenchley, J. M. et al. CD4+ T cell depletion during all stages of HIV disease occurs predominantly in the gastrointestinal tract. J. Exp. Med. 200, 749–759 (2004)
Bleul, C. C., Wu, L., Hoxie, J. A., Springer, T. A. & Mackay, C. R. The HIV coreceptors CXCR4 and CCR5 are differentially expressed and regulated on human T lymphocytes. Proc. Natl Acad. Sci. USA 94, 1925–1930 (1997)
Gondois-Rey, F. et al. Segregation of R5 and X4 HIV-1 variants to memory T cell subsets differentially expressing CD62L in ex vivo infected human lymphoid tissue. AIDS 16, 1245–1249 (2002)
Penn, M. L., Grivel, J. C., Schramm, B., Goldsmith, M. A. & Margolis, L. CXCR4 utilization is sufficient to trigger CD4+ T cell depletion in HIV-1-infected human lymphoid tissue. Proc. Natl Acad. Sci. USA 96, 663–668 (1999)
Cho, Y. S. et al. Phosphorylation-driven assembly of the RIP1-RIP3 complex regulates programmed necrosis and virus-induced inflammation. Cell 137, 1112–1123 (2009)
Pitha, P. M. Innate antiviral response: role in HIV-1 infection. Viruses 3, 1179–1203 (2011)
Samuel, C. E. Antiviral actions of interferons. Clinical Microbiology Reviews 14, 778–809 (2001)
Lamkanfi, M. & Dixit, V. M. The inflammasomes. PLoS Pathog. 5, e1000510 (2009)
Coll, R. C. & O’Neill, L. A. The cytokine release inhibitory drug CRID3 targets ASC oligomerisation in the NLRP3 and AIM2 inflammasomes. PLoS ONE 6, e29539 (2011)
Juliana, C. et al. Anti-inflammatory compounds parthenolide and Bay 11-7082 are direct inhibitors of the inflammasome. J. Biol. Chem. 285, 9792–9802 (2010)
Lamkanfi, M. et al. Glyburide inhibits the Cryopyrin/Nalp3 inflammasome. J. Cell Biol. 187, 61–70 (2009)
MacDonald, K. P. et al. Characterization of human blood dendritic cell subsets. Blood 100, 4512–4520 (2002)
Merad, M., Sathe, P., Helft, J., Miller, J. & Mortha, A. The dendritic cell lineage: ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu. Rev. Immunol. 31, 563–604 (2013)
Descours, B. et al. SAMHD1 restricts HIV-1 reverse transcription in quiescent CD4+ T-cells. Retrovirology 9, 87 (2012)
Acknowledgements
We thank D. N. Levy for the NLENG1 plasmid; L. A. J. O’Neill for CRID3 and parthenolide; R. Collman for the HIV-1 89.6 clone; and Vertex Pharmaceuticals for the VX-765 and VRT-043198 compounds. HIV-infected lymph node tissue was obtained from the SCOPE cohort at HIV/AIDS clinic of the San Francisco General Hospital (SFGH) Positive Health Program, with the help of R. Hoh, and M. Kerbelski. We thank W. Schecter for surgical removal of the lymph nodes from HIV-infected subjects. We thank L. Napolitano and Y. Lie from Monogram Biosciences for performing Trofile assays to determine HIV co-receptor tropism in samples of HIV-infected volunteers. The following reagents were obtained through the AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH: AMD3100, efavirenz and raltegravir. We thank C. Miller, director of the Gladstone Histology Core for performing the immunostaining assays and M. Cavrois, M. Gesner, and J. Tawney for assistance with flow cytometry. We also thank G. Howard and A. L. Lucido for editorial assistance; J. C. W. Carroll, G. Maki, and T. Roberts for graphics arts; and R. Givens and S. Cammack for administrative assistance. Special thanks to N. Roan for comments on the manuscript and to J. Neidleman for stimulating discussions and technical advice. We thank the NIH/NIAID for funding (R21AI102782, 1DP1036502, U19 AI0961133). Funding was also provided by the UCSF/Robert John Sabo Trust Award (G.D.) and A.P. Giannini Foundation Postdoctoral Research Fellowship (K.M.M.). We also acknowledge support from NIH P30 AI027763 (UCSF-GIVI Center for AIDS Research) for support to S.S. and Z.Y., and for Immunology Core services.
Author information
Authors and Affiliations
Contributions
G.D. identified the involvement of caspase 1 and pyroptosis in lymphoid CD4 T-cell death by HIV-1, developed and designed most of the studies, collected the data and wrote the manuscript; N.L.K.G. performed IL-1β protein assays and examined VX-765 in HIV-infected tonsils; X.G. performed FLICA and shRNA analyses in HLACs; Z.Y. analysed caspase cleavage in HIV-infected cultures; K.M.M. examined caspase inhibitors and LDH release assays; O.Z. tested caspase inhibitors, type-I IFN, and pro-IL-1β expression; P.W.H. and H.H. provided HIV-infected lymphoid node from surgeries of SCOPE cohort patients at HIV/AIDS clinic of the San Francisco General Hospital (SFGH); I.M.-A. provided reagents and tissues; S.S. coordinated lymph node biopsies; W.C.G. supervised all of these studies and participated in the preparation of the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
Extended Data Figure 1 Extensive caspase 1 activation in dying lymphoid CD4 T cells infected with either NL4-3 or a primary HIV-1 isolate.
a, Dying CD4 T cells activate caspase 1. HLACs were infected with NL4-3 or with a primary HIV-1 isolate 89.6 obtained from a mixed PBMC culture from an AIDS patient. The 89.6 viral isolate replicates to high titres in primary human cells such as macrophages and lymphocytes. It is highly cytopathic and utilizes both CCR5 and CXCR4 as co-receptors (dual-tropic)18,54. Infected cells were treated either with no drugs or with AMD3100 (250 nM) entry inhibitor, as indicated. Caspase 1 activity was determined by flow cytometry using FLICA 12 h after treatment with nigericin (10 μM) or 3 days after infection with HIV. Notably, equivalent levels of caspase 1 activation were observed in CD4 T cells infected with NL4-3 or 89.6 HIV-1 isolate. AMD3100 prevented caspase 1 activity with both viruses, indicating the abundant presence of CXCR4-expressing target CD4 T cells in these cultures. b, Low levels of caspase 3 activity in dying CD4 T cells. The same cultures as in (a) were tested for caspase 3 activity using FLICA. Compared to caspase 1, infections with NL4-3 and 89.6 HIV-1 isolate induced low levels of caspase 3 activation in dying CD4 T cells. No caspase 3 activation was observed in cells treated with nigericin, which signals the NLRP3 inflammasome to activate caspase 1 (ref. 19), indicating a specific recognition of caspase 1 and caspase 3 activity by the FLICA probes. These data are the representative results of four independent experiments performed in tonsil cells isolated from four different donors.
Extended Data Figure 2 Resting CD4 T cells from tonsil include both naive and memory subsets.
a, CD4 T lymphocytes in lymphoid tissues contain a large population of central memory cells. To identify the sub-populations of CD4 T cells in human tonsil histocultures, we examined the expression pattern of CCR5, CD45RA, CD45RO, CD62L and CD27. Central memory CD4 T cells (TCM) are characterized by expression of CD45RO+/CD62L+ or CD45RO+/CD27+ 29,31,55,56. TCM lack effector function and constantly travel through the lymph nodes in large quantities for antigen sampling, whereas effector memory cell (TEM) mainly migrate to peripheral tissues29,30,31. Analysis of these surface markers revealed at least three distinct maturation phenotypes. The majority of CD4 T lymphocytes exhibit a memory phenotype as determined by surface expression of CD45RO, among them more than two-thirds were found to be central memory cells (CD45RO+/CD62L+ and CD45RO+/CD27+). Similarly, a large population of CCR5-expressing CD4 T cells was found to have central memory phenotype (CCR5+/CD62L+ and CCR5+/CD27+). These findings are in accordance with previous studies in primary human lymphoid cultures12,57,58. b, c, Memory lymphoid CD4 T cells represent preferential targets for productive infection by both the R5- and X4-tropic strains of HIV-1. To determine whether cell maturation influences susceptibility for productive infection, we measured the levels of productive infection using GFP reporter viruses harbouring either an X4-tropic or R5-tropic Env of HIV-1. Except for their select V3 loop envelope determinants, both reporters were derived from the same bicistronic Nef-IRES-GFP clone which produces fully replication-competent viruses16. Interestingly, productive infection of both X4-tropic or R5-tropic viral strains was detected in CXCR4-expressing cells, indicating that the CXCR4 co-receptor is equally present on CCR5-expressing cells, as was previously shown12,57,58,59. Memory CD4 T cells (CD45RO+) were selectively productively infected in cultures infected with either X4-tropic or R5-tropic reporter virus. Similar findings were found in infected cultures activated with CD3/CD38 beads to achieve higher rates of infection. Among the memory CD4 T cells, TEM cells became productively infected in higher quantities than TCM (not shown). These data are the representative results of six independent analyses performed in tonsil cells isolated from six different donors.
Extended Data Figure 3 Necrostatin-1 does not prevent lymphoid CD4 T-cell death and cell lysis in HIV-infected cultures.
a, b, Necrostatin was tested at 1 or 5 μM, a concentration that yields maximal inhibition without inducing toxicity (not shown). Pyroptosis shares cell death features with necroptosis which similarly leads to the release of intracellular contents into the extracellular space9. To test whether cell death involves necrotic signalling, we treated HIV-infected CD4 T cells with necrostatin, a specific inhibitor of RIP1, whose kinase activity is essential for programmed necroptosis to occur60. Concentrations of necrostatin that block necroptotic signalling (not shown) did not inhibit CD4 T-cell depletion in HIV-infected cultures (a), and did not prevent the release of intracellular contents into the culture medium, as indicated by LDH activity in the supernatants (b). Thus, although pyroptosis shares features with necroptosis, these data demonstrate that the signalling pathways linking caspase 1 activation to CD4 T-cell death are specific. Together, these findings indicate that the CD4 T-cell depletion and release of cytoplasmic contents in HIV-infected lymphoid cultures reflects pyroptosis rather than apoptosis or necroptosis. Error bars represent s.e.m. of at least three independent experiments using tonsil cells from at least three different donors. c, Caspase 1 inhibitors prevent CD4 T-cell death in HIV-infected splenic tissues. Splenic HLACs were cultured with no virus or were infected with HIV-1. The HIV-infected cultures were treated as indicated, either with no drugs, efavirenz (100 nM), the caspase 1 inhibitor Ac-YVAD-CMK (50 μM), or the caspases-3 inhibitor Z-DEVD-FMK (50 μM). After 4 days, viable CD4 T cells were counted by flow cytometry. Viable CD4 T cells are presented as the percentage remaining live CD4 T cells using CD8 T cells to normalize each HIV-infected or uninfected culture. Error bars represent s.e.m. from four independent experiments using tonsil cells isolated from four different donors.
Extended Data Figure 4 Induction of type-I interferon is not required to trigger a death response in HIV-infected lymphoid CD4 T cells.
HIV-1 infections induce type-I interferon in vitro and in vivo61. To test the involvement of this antiviral response in modulating CD4 T-cell death, isolated CD4 T cells were infected with HIV-1 in the presence of neutralizing antibodies against the human interferon alpha receptor (IFNAR2), which blocks biological action of type I interferons. To determine the state of interferon signalling, cells were analysed in parallel for the presence of tyrosine-phosphorylated STAT1, which plays a central role in mediating type-I IFN-dependent biological responses, including induction of an antiviral state62. Phosphorylated STAT1 readily appeared in HIV-infected CD4 T cells, but not in HIV-infected cells treated with efavirenz (100 nM), AMD3100 (250 nM) or anti-IFNAR2 neutralizing antibodies (1–5 μg ml−1). Notably, blocking interferon signalling with anti-IFNAR2 neutralizing antibodies did not prevent the death of CD4 T cells by HIV-1, although tyrosine phosphorylation of STAT1 was inhibited indicating effectiveness of the antibody blockade. The data suggest that this antiviral IFN induction is not critical to the onset of the innate immune death response leading to caspase 1 activation and pyroptosis. Error bars represent s.e.m. from three independent experiments using tonsil cells from three different donors.
Extended Data Figure 5 Lymphoid CD4 T cells express detectable levels of ASC and NLRP3 relative to blood-derived monocytes.
The bipartite adaptor protein ASC (PYCARD) plays a central role in the interaction between (NOD)-like receptor and caspase 1 in inflammasome complexes63. Lymphoid CD4 T cells are primed to mount such inflammatory responses, and constitutively express high levels of cytoplasmic pro-IL-1β, but also ASC and NLRP3, compared to blood-derived monocytes. CD4 T lymphocytes express constitutive levels of NLRP3. In contrast to lymphocytes, monocytes require stimulation with TLR ligands such as LPS to induce NLRP3 expression21. Thus, the release of intracellular 5′-ATP by pyroptotic CD4 T cells may provide a second inflammatory stimulus to induce activation of caspase 1 by the NLRP3 inflammasome in nearby CD4 T cells that are already primed as reflected by their high levels of ASC, NLRP3 and pro-IL-1β expression. Thus, pyroptosis activated initially by HIV may result in cascade of new rounds of pyroptosis in primed CD4 T cells by the repeated release of intracellular ATP in a virus-independent manner. Such an ‘auto-inflammation’ scenario could result in persistent rounds of pyroptosis, chronic inflammation and loss of CD4 T cells even when viral loads are reduced by antiretroviral therapy (ART).
Extended Data Figure 6 Low levels of caspase 3 activation in HIV-infected lymphoid CD4 T-cell cultures.
Although the endogenous levels of pro-caspase 3 and pro-caspase 1 expression are similar in lymphoid CD4 T cells, caspase 3 activation in these cells was markedly less abundant after infection with HIV-1, compared to caspase 1. These data are in accord with our findings using fluorescently labelled inhibitor of caspases (FLICA) probes in cultures infected with a GFP reporter HIV-1. In these cultures, the majority of CD4 T cells were abortively infected and showed activation of intracellular caspase 1. No caspase 1 activity was observed in productively infected cells (Fig. 1b). In sharp contrast, caspase 3 activity in these cultures was markedly less abundant, and specifically occurred in productively infected, but not in non-productively infected cells (Fig. 1c). These data are the representative results of three independent experiments performed in tonsillar CD4 T cells isolated from three different donors.
Extended Data Figure 7 Inhibitors of caspase 1, but not of NLRP3, prevent CD4 T-cell death by HIV-1.
a, Quantitative evaluation of bioactive IL-1β secreted in HIV-infected CD4 T-cell cultures using ELISA. Isolated tonsillar CD4 T cells were left uninfected or infected with HIV in the presence of the indicated drugs. Four days after infection, supernatants were filtered through 0.22-μm filter plates and subjected to IL-1β ELISA analysis. A total of 200 μl of supernatant from 2 million isolated CD4 T cells was used for each condition. The assay was performed as described by the manufacturer’s instructions (R&D Systems). Bioactive IL-1β was detected in supernatants of HIV-infected cultures, at levels comparable to those in uninfected cells treated with nigericin. Treatments of HIV-infected cultures with viral or caspase 1 inhibitors, but not caspase 3 inhibitor, reduced accumulation of IL-1β in the supernatants to levels comparable to those detected in uninfected cultures. These finding demonstrate that caspase 1 activation is specifically required for the release of bioactive IL-1β in lymphoid CD4 T cells infected with HIV-1. Error bars represent s.e.m. of three independent experiments using tonsil cells from at least three different donors. b, Inhibitors of caspase 1 and the NLRP3 inflammasome prevent release of mature IL-1β induced by nigericin, but not CD4 T-cell death by HIV-1. Because nigericin engages the NLRP3 inflammasome to activate caspase 1 in lymphoid CD4 T cells, we sought to determine if NLRP3 also similarly controls caspase 1 activity in response to HIV-1 infection. Cell cultures were treated with four separate NLRP3 inhibitors including CRID364, parthenolide65, and the sulfonylureas glyburide66 and glimepiride. Treatments with CRID3, parthenolide or sulfonylureas (not shown) completely inhibited NLRP3-dependent release of mature IL-1β by nigericin, but had no effect on IL-1β release triggered by HIV infection of lymphoid CD4 T-cell cultures (Fig. 3f). c, Treatments with CRID3, parthenolide or sulfonylureas did not prevent HIV-1-mediated CD4 T-cell death. These results suggest that the NLRP3 inflammasome does not control the caspase-1-mediated death responses in lymphoid CD4 T cells abortively infected with HIV-1. Cell death results are represented as ratios of viable CD4 versus CD8 T cells in each HIV-infected or uninfected culture. Error bars represent s.e.m. of four independent experiments using tonsil cells from four different donors.
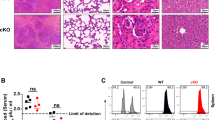
Extended Data Figure 8 Distinct regions of caspase 1 and caspase 3 activity in lymph node of a chronically infected HIV patient.
Inguinal lymph node was collected from a 41-year-old African-American male, infected with an R5-tropic strain of HIV-1. The patient had been on intermittent anti-retroviral therapy between 2004–2009, and stopped anti-retroviral therapy in 2009. This individual exhibited a viral load of 30,173 RNA copies per ml, and CD4 T-cell count of 259 cells per μl. The fresh specimen was immediately subjected to immunostaining in parallel with fresh uninfected human tonsil. Note the immunostain against CD3 highlights the paracortical region, which is almost entirely composed of resting T cells. Note also the sparse presence of CD3-positive T cells in the mantle zones and germinal centres, where lymphocytes become activated (Ki-67) and differentiate into memory and plasma cells. These CD4 T cells are responsible for antigen-dependent activation of B cells in the follicle. Staining for CD11c reveals scattered dendritic cells67,68 in the germinal centre and largely in the mantle zone. HIV p24 Gag expression is located between the mantle zone and germinal centres, where activated CD4 T cells reside. Remarkably, caspase 3 activity also occurs in this anatomical region, which is separated from the majority of non-productively infected T cells in the paracortical zone and exhibit caspase 1 activation, IL-1β processing and pyroptosis. The anti-caspase 1 antibody was raised against a peptide mapping to the C terminus of caspase 1 p20 of human origin and detects both the cleaved p20 subunit and the precursor of caspase 1. Therefore, in the absence of an equivalent uninfected lymph node it is hard to absolutely determine whether abortive HIV-1 infection affects pro-caspase expression. However, staining of uninfected tonsil or spleen (not shown) tissue revealed no positive HIV p24 Gag, active or pro-caspase 1, bioactive IL-1β or annexin V signals. These data closely correlate with the findings in HIV-infected HLACs where the 95% of the CD4 T cells are non-productively infected CD4 T cells and show activation of intracellular caspase 1, whereas caspase 3 activity is markedly less abundant and specifically occurs in productively infected CD4 T cells. GC, germinal centre; MZ, mantle zone; PC, paracortical zone.
Extended Data Figure 9 Targeting caspase 1 via an orally bioavailable small molecule inhibitor prevents lymphoid CD4 T-cell death by HIV-1.
a, VX-765 is a cell permeable pro-drug (1) that requires intracellular esterase cleavage in the cell to yield the aldehyde functionality (green) of the drug VRT-043298 (2b), which acts as a potent caspase 1 inhibitor. Adapted from ref. 38 with permission. b, VX-765 prevents CD4 T-cell death in a dose-dependent manner in HIV-infected lymphoid tissues. HLACs were either not infected or infected with HIV-1 in the absence of drugs or in the presence of efavirenz (100 nM), AMD3100 (250 nM) or VX-765 (0.05, 0.5 or 5 μM) as indicated. VX-765 was added to the cultures 4 hours before infection to allow absorption and processing by the cells. Flow cytometry plots depict gating on live cells based on the forward-scatter versus side-scatter profile of the complete culture. These results are representative of three independent experiments performed using tonsil cells from three different donors.
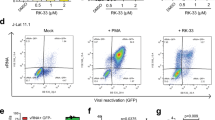
Extended Data Figure 10 Treatment with a caspase1 inhibitor does not increase productive HIV-1 infection.
To determine whether inhibition of caspase-1-mediated pyroptosis would result in higher levels of productive HIV-1 infection, tonsillar HLACs were treated with AMD3100 or with the caspase 1 inhibitor Ac-YVAD-CMK before infection with a GFP reporter virus (NLENG1). After 5 days, flow cytometry analysis of the infected cultures revealed no increase in GFP-positive cells in the infected cultures treated with the caspase 1 inhibitor Ac-YVAD-CMK. This result likely reflects the continued function of the host restriction factor SAMHD1 (refs 35, 69). These findings argue against the possibility that pyroptosis functions as a defence against productive infection. Instead, pyroptosis appears to represent an overall harmful response that centrally contributes to HIV pathogenesis. These results also argue that interdiction of the pyroptosis pathway with caspase 1 inhibitors would produce beneficial rather than harmful therapeutic effects.
Rights and permissions
About this article
Cite this article
Doitsh, G., Galloway, N., Geng, X. et al. Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection. Nature 505, 509–514 (2014). https://doi.org/10.1038/nature12940
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature12940
This article is cited by
-
Mitochondrial disturbance related to increased caspase-1 of CD4+T cells in HIV-1 infection
BMC Infectious Diseases (2024)
-
Epigenetic modulation of myeloid cell functions in HIV and SARS-CoV-2 infection
Molecular Biology Reports (2024)
-
Knockdown of ELF4 aggravates renal injury in ischemia/reperfusion mice through promotion of pyroptosis, inflammation, oxidative stress, and endoplasmic reticulum stress
BMC Molecular and Cell Biology (2023)
-
Gasdermin D-mediated pyroptosis is regulated by AMPK-mediated phosphorylation in tumor cells
Cell Death & Disease (2023)
-
Inflammasome activation occurs in CD4+ and CD8+ T cells during graft-versus-host disease
Cell Death & Disease (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
