
Abstract
In the mammalian cerebral cortex the diversity of interneuronal subtypes underlies a division of labour subserving distinct modes of inhibitory control1,2,3,4,5,6,7. A unique mode of inhibitory control may be provided by inhibitory neurons that specifically suppress the firing of other inhibitory neurons. Such disinhibition could lead to the selective amplification of local processing and serve the important computational functions of gating and gain modulation8,9. Although several interneuron populations are known to target other interneurons to varying degrees10,11,12,13,14,15, little is known about interneurons specializing in disinhibition and their in vivo function. Here we show that a class of interneurons that express vasoactive intestinal polypeptide (VIP) mediates disinhibitory control in multiple areas of neocortex and is recruited by reinforcement signals. By combining optogenetic activation with single-cell recordings, we examined the functional role of VIP interneurons in awake mice, and investigated the underlying circuit mechanisms in vitro in auditory and medial prefrontal cortices. We identified a basic disinhibitory circuit module in which activation of VIP interneurons transiently suppresses primarily somatostatin- and a fraction of parvalbumin-expressing inhibitory interneurons that specialize in the control of the input and output of principal cells, respectively3,6,16,17. During the performance of an auditory discrimination task, reinforcement signals (reward and punishment) strongly and uniformly activated VIP neurons in auditory cortex, and in turn VIP recruitment increased the gain of a functional subpopulation of principal neurons. These results reveal a specific cell type and microcircuit underlying disinhibitory control in cortex and demonstrate that it is activated under specific behavioural conditions.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Change history
30 October 2013
Figure 1 in the HTML version was corrupted. This has now been updated.
27 November 2013
Figure 3b and its legend were corrected and the number 98 was changed to 97 in three places on page 2.
References
Silberberg, G. & Markram, H. Disynaptic inhibition between neocortical pyramidal cells mediated by Martinotti cells. Neuron 53, 735–746 (2007)
Kapfer, C., Glickfeld, L. L., Atallah, B. V. & Scanziani, M. Supralinear increase of recurrent inhibition during sparse activity in the somatosensory cortex. Nature Neurosci. 10, 743–753 (2007)
Gentet, L. J. et al. Unique functional properties of somatostatin-expressing GABAergic neurons in mouse barrel cortex. Nature Neurosci. 15, 607–612 (2012)
Cardin, J. A. et al. Driving fast-spiking cells induces gamma rhythm and controls sensory responses. Nature 459, 663–667 (2009)
Sohal, V. S., Zhang, F., Yizhar, O. & Deisseroth, K. Parvalbumin neurons and gamma rhythms enhance cortical circuit performance. Nature 459, 698–702 (2009)
Isaacson, J. S. & Scanziani, M. How inhibition shapes cortical activity. Neuron 72, 231–243 (2011)
Ascoli, G. A. et al. Petilla terminology: nomenclature of features of GABAergic interneurons of the cerebral cortex. Nature Rev. Neurosci. 9, 557–568 (2008)
Carandini, M., Heeger, D. J. & Movshon, J. A. Linearity and normalization in simple cells of the macaque primary visual cortex. J. Neurosci. 17, 8621–8644 (1997)
Salinas, E. & Thier, P. Gain modulation: a major computational principle of the central nervous system. Neuron 27, 15–21 (2000)
Acsády, L., Gorcs, T. J. & Freund, T. F. Different populations of vasoactive intestinal polypeptide-immunoreactive interneurons are specialized to control pyramidal cells or interneurons in the hippocampus. Neuroscience 73, 317–334 (1996)
Dávid, C., Schleicher, A., Zuschratter, W. & Staiger, J. F. The innervation of parvalbumin-containing interneurons by VIP-immunopositive interneurons in the primary somatosensory cortex of the adult rat. Eur. J. Neurosci. 25, 2329–2340 (2007)
Hájos, N., Acsady, L. & Freund, T. F. Target selectivity and neurochemical characteristics of VIP-immunoreactive interneurons in the rat dentate gyrus. Eur. J. Neurosci. 8, 1415–1431 (1996)
Jiang, X., Wang, G., Lee, A. J., Stornetta, R. L. & Zhu, J. J. The organization of two new cortical interneuronal circuits. Nature Neurosci. 16, 210–218 (2013)
Xu, H., Jeong, H. Y., Tremblay, R. & Rudy, B. Neocortical somatostatin-expressing GABAergic interneurons disinhibit the thalamorecipient layer 4. Neuron 77, 155–167 (2013)
Pfeffer, C. K., Xue, M., He, M., Huang, Z. J. & Scanziani, M. Inhibition of inhibition in visual cortex: the logic of connections between molecularly distinct interneurons. Nature Neurosci. 16, 1068–1076 (2013)
Lovett-Barron, M. et al. Regulation of neuronal input transformations by tunable dendritic inhibition. Nature Neurosci. 15, 423–430 (2012)
Lee, S. H. et al. Activation of specific interneurons improves V1 feature selectivity and visual perception. Nature 488, 379–383 (2012)
Letzkus, J. J. et al. A disinhibitory microcircuit for associative fear learning in the auditory cortex. Nature 480, 331–335 (2011)
Xu, X., Roby, K. D. & Callaway, E. M. Immunochemical characterization of inhibitory mouse cortical neurons: three chemically distinct classes of inhibitory cells. J. Comp. Neurol. 518, 389–404 (2010)
Rudy, B., Fishell, G., Lee, S. & Hjerling-Leffler, J. Three groups of interneurons account for nearly 100% of neocortical GABAergic neurons. Dev. Neurobiol. 71, 45–61 (2011)
Taniguchi, H. et al. A resource of Cre driver lines for genetic targeting of GABAergic neurons in cerebral cortex. Neuron 71, 995–1013 (2011)
Madisen, L. et al. A toolbox of Cre-dependent optogenetic transgenic mice for light-induced activation and silencing. Nature Neurosci. 15, 793–802 (2012)
Csicsvari, J., Hirase, H., Czurko, A. & Buzsaki, G. Reliability and state dependence of pyramidal cell-interneuron synapses in the hippocampus: an ensemble approach in the behaving rat. Neuron 21, 179–189 (1998)
Oliva, A. A., Jr, Jiang, M., Lam, T., Smith, K. L. & Swann, J. W. Novel hippocampal interneuronal subtypes identified using transgenic mice that express green fluorescent protein in GABAergic interneurons. J. Neurosci. 20, 3354–3368 (2000)
Chattopadhyaya, B. et al. Experience and activity-dependent maturation of perisomatic GABAergic innervation in primary visual cortex during a postnatal critical period. J. Neurosci. 24, 9598–9611 (2004)
Williford, T. & Maunsell, J. H. Effects of spatial attention on contrast response functions in macaque area V4. J. Neurophysiol. 96, 40–54 (2006)
Paspalas, C. D. & Papadopoulos, G. C. Serotoninergic afferents preferentially innervate distinct subclasses of peptidergic interneurons in the rat visual cortex. Brain Res. 891, 158–167 (2001)
Kawaguchi, Y. Selective cholinergic modulation of cortical GABAergic cell subtypes. J. Neurophysiol. 78, 1743–1747 (1997)
Arroyo, S., Bennett, C., Aziz, D., Brown, S. P. & Hestrin, S. Prolonged disynaptic inhibition in the cortex mediated by slow, non-alpha7 nicotinic excitation of a specific subset of cortical interneurons. J. Neurosci. 32, 3859–3864 (2012)
Alitto, H. J. & Dan, Y. Cell-type-specific modulation of neocortical activity by basal forebrain input. Front Syst Neurosci 6, 79 (2013)
Zhang, F. et al. The microbial opsin family of optogenetic tools. Cell 147, 1446–1457 (2011)
Kvitsiani, D. et al. Distinct behavioural and network correlates of two interneuron types in prefrontal cortex. Nature 498, 363–366 (2013)
Schmitzer-Torbert, N., Jackson, J., Henze, D., Harris, K. & Redish, A. D. Quantitative measures of cluster quality for use in extracellular recordings. Neuroscience 131, 1–11 (2005)
Schwarz, G. E. Estimating the dimension of a model. Ann. Stat. 6, 461–464 (1978)
Lorincz, A. & Nusser, Z. Cell-type-dependent molecular composition of the axon initial segment. J. Neurosci. 28, 14329–14340 (2008)
Katona, I. et al. Molecular composition of the endocannabinoid system at glutamatergic synapses. J. Neurosci. 26, 5628–5637 (2006)
Varga, V. et al. The presence of pacemaker HCN channels identifies theta rhythmic GABAergic neurons in the medial septum. J. Physiol. 586, 3893–3915 (2008)
Acknowledgements
We are grateful to B. Mensh, S. Ranade, N. Spruston and A. M. Zador for comments and discussions; S. Ranade and R. Eifert for assistance for microdrive design; S. G. Koh, A. Reid, H. Li and Y. Kim for help with experimental setup; A. M. Zador for use of in vitro electrophysiology equipment; B. Burbach for technical assistance; and J. Kuhl for help with figures. This research was supported by grants from NIH NINDS R01NS075531, the Klingenstein, John Merck, and Sloan Foundations to A.K. and from NIH NIMH U01MH078844 to Z.J.H. B.H. received support from the Swartz Foundation and Marie Curie International Outgoing Fellowship within the EU Seventh Framework Programme for Research and Technological Development. D.K. received support from The Robert Lee and Clara Guthrie Patterson Trust Postdoctoral Fellowship and Human Frontier Science Program.
Author information
Authors and Affiliations
Contributions
H.-J.P., B.H. and A.K. designed the experiments. H.-J.P. and B.H. performed the experiments and analysed data. D.K. set up in vivo optogenetics-assisted recordings. J.I.S. designed custom behaviour and stimulation systems for the behavioural task. Z.J.H. provided the VIP-IRES-Cre mouse line. H.-J.P., B.H. and A.K. wrote the manuscript with comments from Z.J.H., J.S. and D.K.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Extended data figures and tables
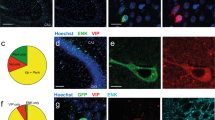
Extended Data Figure 1 Specificity and efficiency of ChR2 expression, recording locations and layer 1 VIP neurons.
a, VIP-Cre:Ai32 (ACx). b, VIP-Cre::AAV-DIO-ChR2 (mPFC). Note that some green puncta were not somata and only somata were used for quantification. c, Quantification of the overlap of ChR2–YFP (green) with anti-VIP (red) in ACx. Overlap = 88 ± 6.6% (49 of 54 neurons), 6 slices from 4 mice (see Methods for a note on caveats). d, Quantification of the overlap of ChR2–YFP (green) with anti-VIP (red) in mPFC. Overlap = 97 ± 3.7% (35 of 36 neurons), 5 slices from 4 mice. Scale bar, 50 μm. e, f, Recording location in mPFC and ACx. Microdrives accommodating 6 tetrodes and 1 optical fibre were implanted in the ACx (e) or the mPFC (f). Recording sites were confirmed by histology using DiI (red) that was applied to the optical fibre before implantation. Histology results showed that the electrode locations were biased towards the middle layers. Green, VIP neurons expressing ChR2–YFP; red, DiI. Scale bar, 200 μm. g, h, Most VIP neurons were located in layer 2/3, with a smaller fraction in layer 1. VIP comprised about 10% of layer 1 neurons. g, VIP neuron in layer 1 of the ACx (arrow; 6 VIP/60 layer 1 ACx neurons, n = 6 slices from 2 mice). h, VIP neuron in layer 1 of the mPFC (5 VIP/56 layer 1 mPFC neurons, n = 6 slices from 2 mice). Green, VIP; red, NeuN (neuronal marker) staining. Scale bar, 100 μm.
Extended Data Figure 2 Three distinct populations responsive to photostimulation in mPFC and spike width versus firing rate.
a–c, Raster plots and PSTHs aligned to photostimulation for three distinct populations in mPFC. Examples of a directly activated (VIP, a), an inhibited (b) and a delayed activated neuron (c). Stimulation frequency, 10–20 Hz. d, Photostimulation-evoked spike probability of a VIP interneuron. Left: raster plot. Right: firing probability as a function of photostimulation frequency. When all light pulses were considered, spike probability decreased with stimulation frequency (dark blue). However, the first 5 light pulses reliably evoke action potentials up to 100 Hz (light blue; spike probability = 0.78 at 100 Hz). e, Relative light-induced firing rate change (log scale) versus latency of the maximal effect (peak/trough of PSTH). Three separate groups are apparent in mPFC: short-latency activated (VIP, green), inhibited (purple) and delayed activated (light brown). f, g, Top left: baseline firing rate versus spike width in ACx (f) and mPFC (g). Top right: cumulative fraction of firing rate. Bottom: cumulative fraction of spike width. Green, directly activated (VIP); purple, inhibited; light brown, delayed activated group; dark grey, unidentified neurons. Light grey depicts neurons for which inhibition could not reliably be assessed because of very low baseline firing rates (see Methods).
Extended Data Figure 3 Inhibited and activated (Inh-act) population is a subgroup of the inhibited neurons.
a, Average PSTH aligned to photostimulation (1-ms pulses) for Inh-act cells (a subgroup of the inhibited neurons, Figs 1 and 3). The colour code of Fig. 1g, h applies. Inh-act neurons (purple) show initial inhibition followed by delayed activation after 1-ms pulses. b, Top: example raster plot aligned to auditory stimuli of a tone-responsive Inh-act neuron. Shading indicates the stimulation windows (grey, tone only; blue, tone- and photostimulation). Dashed boxes indicate time windows for frequency tuning analysis (early, 0–25 ms; late, 75–100 ms). Bottom: frequency tuning curves of tone-responsive Inh-act neurons (population average, n = 14). Bottom left: tuning curve for the early time window (0–25 ms). Bottom right: tuning curve for the late time window (75–100 ms). Simultaneous photostimulation (100 ms) decreased the tone-evoked firing rates of Inh-act neurons in the early time window, whereas it increased the firing rates in the late time window. This pattern resembled the inhibition-activation sequence elicited by the 1 ms light pulses (a).
Extended Data Figure 4 Light-intensity-dependent changes in spike probability, delay and jitter in VIP interneurons in vitro.
a, Example traces of action potentials evoked by different light intensities. Blue bar, light stimulation. Scale bar, 10 ms, 10 mV. b–d, Quantification. Spike probability (b) increased whereas delay (c) and jitter (d) decreased with increasing light intensities. The highest two intensities were used in the in vitro experiments.
Extended Data Figure 5 Photostimulation of VIP increases c-Fos in pyramidal neurons in mPFC.
a, Schematic of c-Fos experiment. Animals were anaesthetized for 2 h to reduce the background c-Fos levels and photostimulation was applied. The expression level of c-Fos was captured 1 h after photostimulation. b, Representative images of different experimental conditions. CTRL1, no ChR2 expression with photostimulation; CTRL2, ChR2 expression without photostimulation; EXP, ChR2 expression with photostimulation. Left column: green, expression of ChR2.YFP; middle: white, c-Fos staining; right: merged images. Scale bar, 200 μm. c, Quantification of c-Fos levels. CTRL1, n = 64 c-Fos immunopositive neurons, n = 4 mice; CTRL2, n = 58 neurons, n = 4 mice; EXP, n = 252 neurons, n = 4 mice. d–f, Representative images from different experimental conditions. White, c-Fos staining; green, ChR2 expression; red, CaMKIIα staining. The arrow indicates a c-Fos-immunopositive VIP neuron. Scale bar, 50 μm. Note that some overlapping signals (c-Fos and CaMKIIα) are hard to appreciate in this image due to low resolution and uneven immunostaining. Additionally, owing to the different signal strength, CaMKIIα immunopositivity is hard to appreciate for neurons that are slightly above or below the focal plane, whereas the strong c-Fos immunoreactivity is still detectable. For this reason, additional high-power images were presented in i. g, Co-localization of markers with c-Fos. h, Fraction of c-Fos-positive cells. Among CaMKIIα-positive neurons, the proportion of c-Fos-immunopositive cells was significantly higher in the experimental group as compared with the controls. CTRL1, fraction = 11/208 (c-Fos/CaMKIIα), 4 mice; CTRL2, fraction = 3/107, 3 mice; EXP, fraction = 66/257, 4 mice. Approximately 64% (7 of 11) of the ChR2-expressing neurons were c-Fos-immunopositive. i, High-resolution images of the co-localization between c-Fos and CaMKIIα in f. Owing to low resolution and uneven staining of CaMKIIα, some c-Fos signals seemingly do not co-localize with CaMKIIα-positive neurons. However, in high-resolution images, the co-localization is clearer. Top: example of a weakly stained CaMKIIα-positive neuron (arrow). In the high-resolution image, CaMKIIα staining is apparent. Bottom: attributed to differences in immunofluorescence strength between c-Fos and CaMKIIα, neurons slightly out of focus may appear c-Fos-positive and CaMKIIα-negative. However, when the focal plane was adjusted, the co-localization became apparent. Blue, c-Fos; green, ChR2–YFP; red, CaMKIIα.
Extended Data Figure 6 Responses of VIP neurons in ACx during auditory stimulation, alone or combined with photostimulation.
a, Raster plot (left) and PSTH (right) aligned to the onset of combined tone and photostimulation. Although all VIP neurons were responsive to photostimulation, individual VIP neurons showed heterogeneous response profiles. One VIP neuron (top) showed accommodation during the 200-ms stimulation; two neurons (second and fourth) showed transient response; one neuron (third) fired persistently throughout the stimulation. Shaded boxes (left) or coloured lines (right) indicate the stimulation duration. b, Average frequency tuning curve of VIP neurons (n = 4). All 4 VIP neurons in the ACx for which tuning curves were recorded were responsive to pure tones; however, their tuning properties showed considerable heterogeneity.
Extended Data Figure 7 Auditory response profiles of different neuronal groups.
a, b, Single-cell examples (top, raster plot; middle, PSTH) and population average (bottom, PSTH) of responses evoked by a brief 1-ms light pulse. a, Delayed activated and tone-responsive neurons. b, Delayed activated and tone-unresponsive neurons. c, d, Single-cell examples (top, raster plot; middle, PSTH) and population average (bottom, PSTH) of responses evoked by combined auditory and light stimulation (100 ms). c, Delayed activated and tone-responsive neurons. d, Delayed activated and tone-unresponsive neurons. Grey shaded box, tone stimulation; blue shaded box, tone plus light stimulation. e, PSTH of tone-responsive (red) and tone-unresponsive (brown) delayed activated neurons for 100-ms light pulses (without auditory stimulation). This experiment was performed in a subset of the frequency tuning experiments. f, Frequency tuning curve of tone-responsive neurons not modulated by 1-ms light pulses. g, Fitting of one-parameter gain control models on tuning curve modulation of inhibited and delayed activated neurons. Tuning curves recorded during photostimulation were fitted with one-parameter models representing the scaled (multiplicative model) or shifted (additive model) versions of the baseline tuning curves (that is, without photostimulation). The ratio of the least squared errors of the two model fits is plotted as a function of relative firing rate change after 1-ms light pulses on a logarithmic scale, for inhibited (purple) and delayed activated (brown) neurons (minus infinity corresponds to complete abolishment of firing). An error ratio >1 corresponds to a better fit of the additive model, whereas <1 means better fit of the multiplicative gain model. See also Methods.
Extended Data Figure 8 VIP neurons strongly respond to punishment in ACx.
a, Raster plots (top) and PETHs (bottom) aligned to reinforcement (reward, green or punishment, red) for all VIP neurons recorded in the auditory go/no-go task. All VIP neurons were strongly recruited by punishment (foot shock: 1–4, marked by asterisks; air puff: 5–10), whereas water reward induced weaker and more sustained activation (9 of 10 cells showed significant firing rate increase for reward, see main text and Methods). b, Raster plots and PETHs of example unidentified neurons. Type 1 neurons (left) tended to be activated by tone onset. Type 2 neurons (right) tended to be inhibited by tone onset. c, Top: normalized average PETH of VIP (green) and concurrently recorded non-VIP neurons aligned to feedback (left) and tone onset (right) for hit (top) and false alarm trials (bottom). VIP neurons showed an abrupt increase of firing rate after punishment (bottom left). The oscillatory pattern of VIP activation around reward delivery is a consequence of rhythmic firing-rate modulations following the highly stereotypic pattern of licking in 4 of 10 VIP neurons (1 of 130 in non-VIP neurons). Grey, feedback-activated unidentified neurons (type 1). Pink, feedback-inhibited unidentified neurons (type 2). Insets, zoomed-in plots of PETHs. Arrows indicate the difference in activation pattern between VIP and unidentified type 1 neurons.
Extended Data Figure 9 Accounting for the temporal difference between VIP-Cre::Ai32 (ACx) and VIP-Cre::AAV.ChR2 (mPFC).
We observed a temporal difference in the firing pattern of VIP neurons between the ChR2 reporter line (Ai32) and the virus-injected (AAV.ChR2) mice. We speculated that this difference could stem from the mutation in ChR2. The mutation in ChR2(H134R) of Ai32 mice produces larger currents and slower kinetics than ChR2(ChETA; AAV.ChR2). As a consequence, VIP neurons in VIP-Cre::Ai32 can fire bursts in response to single 1-ms pulses and the activation can last more than 20 ms (a). This sustained activity of VIP neurons prolonged the temporal dynamics of downstream neurons. a, Examples of VIP neurons that burst to 1-ms photostimulation in the ACx of VIP-Cre::Ai32 mice. b, Model explaining the temporal difference between VIP-Cre::Ai32 and VIP-Cre::AAV-DIO-ChR2(ChETA). In VIP-Cre::Ai32 mice, almost all VIP neurons express ChR2 and exert stronger inhibition on the inhibited neuron group. Because the duration of VIP (green) activation varies, individual inhibited neurons (Inh, purple) receive different degrees of inhibition (strength and duration) from VIP neurons, therefore their firing rates recover to baseline at different time points. This variation propagates to the delayed activated group (dAct, orange), activation of which can start at different time instances.
Rights and permissions
About this article
Cite this article
Pi, HJ., Hangya, B., Kvitsiani, D. et al. Cortical interneurons that specialize in disinhibitory control. Nature 503, 521–524 (2013). https://doi.org/10.1038/nature12676
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature12676
This article is cited by
-
Local origin of excitatory–inhibitory tuning equivalence in a cortical network
Nature Neuroscience (2024)
-
Fear extinction rescuing effects of dopamine and L-DOPA in the ventromedial prefrontal cortex
Translational Psychiatry (2024)
-
Ketamine triggers a switch in excitatory neuronal activity across neocortex
Nature Neuroscience (2023)
-
Adaptive mechanisms facilitate robust performance in noise and in reverberation in an auditory categorization model
Communications Biology (2023)
-
A sleep-active basalocortical pathway crucial for generation and maintenance of chronic pain
Nature Neuroscience (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
