
Abstract
Angiotensin -converting enzyme 2 (ACE2) is a regulator of the renin angiotensin system involved in acute lung failure, cardiovascular functions and severe acute respiratory syndrome (SARS) infections in mammals1,2,3. A gene encoding a homologue to ACE2, termed collectrin (Tmem27), has been identified in immediate proximity to the ace2 locus4. The in vivo function of collectrin was unclear. Here we report that targeted disruption of collectrin in mice results in a severe defect in renal amino acid uptake owing to downregulation of apical amino acid transporters in the kidney. Collectrin associates with multiple apical transporters and defines a novel group of renal amino acid transporters. Expression of collectrin in Xenopus oocytes and Madin–Darby canine kidney (MDCK) cells enhances amino acid transport by the transporter B0AT1. These data identify collectrin as a key regulator of renal amino acid uptake.
Similar content being viewed by others
Main
ACE2 is a chimaeric protein that has emerged from the duplication of two genes: homology with ACE at the catalytic domain and homology with collectrin in the membrane proximal domain (Fig. 1a). In the mouse, collectrin is expressed in the kidneys4 and pancreas5,6 and to a lesser extent in the intestine (duodenum, jejunum and ileum), liver, heart and stomach (Supplementary Fig. 1). To assess the in vivo function of collectrin, we generated collectrin knockout mice (Fig. 1b). Mice were crossed with the transgenic Cre deleter strain to ubiquitously delete collectrin7 (Supplementary Fig. 2a). The null mutation of collectrin was verified by the absence of collectrin messenger RNA transcripts and protein (Supplementary Fig. 2b, c). ACE2 expression in the kidneys was not altered in collectrin mutant mice (Supplementary Fig. 2c). Collectrin null mice were born at the expected mendelian frequency, indistinguishable from their heterozygous and wild-type littermates. Collectrin mutant males (collectrin-/y) and females (collectrin-/-) were fertile and we failed to observe any overt morphological alterations in all organs analysed, including kidney, for up to six months of age (Fig. 1c).

a, Schematic outline of homologous regions between collectrin and ACE2. Collectrin lacks the HEMGH carboxypeptidase domain. The black boxes indicate the transmembrane domains. Leader peptides are shown in light blue. b, Gene targeting (flox-and-delete) strategy for the generation of collectrin mutant alleles. Upon breeding with Cre deleter mice, animals with excision of exon 2 were selected. c, H&E-stained sections of kidney isolated from six-month-old collectrin+/y and collectrin-/y mice. Top panels, overview over cortex; bottom panels, normal medullary structures. d, At 4 °C, large white precipitates form in the urine of collectrin null (-/y) but not control wild-type (+/y) mice. These precipitates disappear upon fasting for 24 h. e, Formation of needle-like crystals in the urine of collectrin null mice. f, HPLC analysis of crystals isolated from the urine of collectrin-/y mice. The crystals co-elute with tyrosine and phenylalanine. mAU, milli-absorbance units. g, Excretion of all amino acids tested in collectrin-/y mice. Urine was collected from four-month-old collectrin+/y and collectrin-/y littermates and analysed by HPLC. Values are mean ± s.e.m. of ten mice per group. P < 0.05 (Student’s t-test and analysis of variance, ANOVA).
Collectrin was originally reported to be expressed in collecting ducts4 and β-islet cells of the pancreas5,6. We therefore assessed whether collectrin is involved in renal salt and/or glucose balance. Na+ excretion in urine was comparable in collectrin+/y and collectrin-/y mice. Loss of collectrin also did not affect the levels of calcium, potassium, phosphate, chloride, urea, uric acid or creatinine in urine and blood (Supplementary Table 1). collectrin mutant mice had the same glucose concentration as their wild-type littermates in serum and urine. Surprisingly, storage of the urine samples collected from mutant mice at 4 °C resulted in the formation of large white precipitates (Fig. 1d). Morphological analysis revealed large needle-like crystals in the urine of the collectrin null mice whereas such crystals were never observed in wild-type mice (Fig. 1e). Biochemically, these crystals were not composed of common urinary crystal components such as ammonium urate, sodium urate, calcium oxalate dihydrate or calcium phosphate. High performance liquid chromatography (HPLC) analyses showed that these crystals contained ∼10% phenylalanine (Phe) and 90% tyrosine (Tyr) (see Fig. 1f). When animals were fed a tyrosine-free and phenylalanine-free diet or when the animals were fasting, no crystals were formed in the urine of collectrin null mice (Fig. 1d), indicating that the observed changes are not caused by tyrosinaemia8,9. Thus, loss of collectrin expression results in the formation of tyrosine/phenylalanine crystals in the urine.
To determine whether collectrin deficiency caused a more general defect in amino acid handling in the kidneys, we assessed urinary amino acid contents. Whereas in wild-type mice almost all amino acids are reabsorbed from the urine, deletion of collectrin resulted in severe leakage of all amino acids (Fig. 1g). Concentrations of the amino acids were also lower in the serum of mutant mice, although amino acids were still present (Supplementary Fig. 3). Urinary output was higher in collectrin-/y mice (3.31 ± 0.25 ml urine per 24 h) as compared to collectrin+/y littermates (1.65 ± 0.28 ml urine per 24 h) which may be owing to the higher concentration of osmotically active solutes in urine from collectrin-deficient mice. Water restriction for 24 h demonstrated that collectrin-deficient mice were not able to concentrate urine to the same degree, confirming that the higher urinary output is due to a primarily renal defect (Supplementary Fig. 4). Thus, collectrin is critical for normal amino acid reabsorption in the kidney.
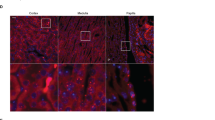
Amino acid reabsorption occurs in the proximal tubules of the kidney10. Immunohistochemical staining showed that collectrin is indeed expressed at the luminal side of brush border membranes in proximal tubules (Fig. 2a), which is a specialized microenvironment involved in amino acid reabsorption11. Biochemically, we confirmed specific expression of collectrin in brush border membrane vesicles (BBMV) isolated from the kidney of collectrin+/y, but not collectrin-/y mice (Fig. 2b). BBMV isolates were free from contamination (assessed by the absence of the collecting duct marker aquaporin 2; Fig. 2b). In addition, collectrin mRNA was found to be highly expressed in proximal tubules (Fig. 2c). Thus, collectrin is localized in brush border membranes of proximal tubules.

a, Immunodetection of collectrin (green) in kidneys of collectrin+/y and collectrin-/y mice. Note that collectrin localizes to proximal tubules. b, Western blot analysis of brush border membranes from collectrin+/y and collectrin-/y mice. Protein extracts from total lysates (50 μg per lane) and renal brush border membranes (10 μg per lane) were analysed using antibodies to collectrin, aquaporin (AQP2), and β-actin. c, Collectrin mRNA expression in proximal tubules (PT) and total kidneys (T). RNA was isolated from wild-type mice and reverse transcribed (+RT). Equal loading was confirmed by Gapdh. d, e, Western blot analysis of proteins (50 μg) from brush border membranes isolated from collectrin+/y and collectrin-/y mice. In d and e, data from three different mice are shown for each genotype. Molecular sizes are indicated in kDa. f, Na+-dependent uptake of the amino acids phenylalanine (Phe) and glutamine (Gln) and uptake of phosphate into BBMVs from collectrin+/y and collectrin-/y mice. All experiments were done in triplicates from each kidney. Values are mean ± s.e.m. of four mice per group. *P < 0.001 (Student’s t-test).
Because collectrin is predicted to be a type I transmembrane protein having only one transmembrane helix4, it was unlikely that collectrin itself forms an amino acid transport pore. Because collectrin expression parallels that of the B0 cluster of the solute carrier SLC6 proteins B0AT1, XT3s1/SIT1, XT2 and XT3, a recently identified subfamily of Na+-dependent transporters of neutral amino acids12,13,14,15,16,17, we assessed whether loss of collectrin may affect these amino acid transporters. Immunoblots of BBMVs revealed that the neutral amino acid transporter B0AT1 was markedly downregulated in collectrin-/y mice (Fig. 2d and Supplementary Fig. 5a, b). Protein expression of the three other members of the SLC6 family XT3s1/SIT1, XT2 and XT3, and expression of EAAC1, the apical transporter responsible for glutamate/aspartate reabsorption18, was also markedly downregulated (Fig. 2e). By contrast, expression of b0,+AT (also known as slc7a9) (Fig. 2e), the exchanger mediating apical uptake of cystine, arginine, lysine and ornithine19, and expression of the basolateral amino acid transporter subunits 4F2hc and LAT2 (ref. 10) remained unchanged (Supplementary Fig. 5b). Expression of the sodium/phosphate co-transporter Na+/Pi IIa was also not affected in collectrin null mice (Fig. 2d). mRNA expression of all these transporters was comparable in collectrin+/y and collectrin-/y mice (Supplementary Fig. 6). Immunoblotting of total kidney membranes demonstrated that the abundance of these amino acid transporters was also decreased to a similar extent as observed in BBMVs (Supplementary Fig. 5b).
To test whether reduced expression resulted in decreased amino acid transport activity, we isolated BBMVs and measured Na+-dependent amino acid uptake rates. As expected from the urinary loss of B0AT1 substrates, the uptake rate of glutamine and phenylalanine in BBMV from kidneys of collectrin null mice was decreased compared to control littermates, whereas uptake of phosphate was not affected (Fig. 2f). These data show that loss of collectrin is associated with reduced expression and impaired activity of apical, Na+-dependent amino acid transporters in the kidney.
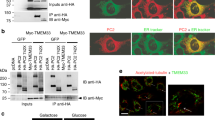
To identify the molecular pathway by which collectrin regulates amino acid transporters, we first examined whether collectrin can associate with B0AT1 and/or other transporters. Immunoprecipitations from BBMV lysates showed direct binding between collectrin and B0AT1, XT2 and XT3 in collectrin+/y, but not collectrin-/y mice (Fig. 3a). Moreover, collectrin co-localized with B0AT1 in the early proximal S1 tubule (Fig. 3b). As a control, collectrin did not associate with b0,+AT (Fig. 3a). The specificity of the collectrin band was confirmed by pre-incubation with the antigenic peptide (Supplementary Fig. 7a, b). Renal amino acid transporters such as LAT2 or y+LAT1 form functional heterodimers with a second subunit, 4F2hc (ref. 10). In all known instances, this heterodimerization is stabilized by disulphide bonds20. Under reducing conditions (+DTT), collectrin appears as a single band with gel mobility of ∼44 kDa, whereas B0AT1 can be detected as a single band that corresponds to a monomeric form of B0AT1 (Fig. 3c). Unlike 4F2hc and its catalytic subunits, B0AT1 and collectrin migrate under non-reducing (no dithiothreitol, -DTT) conditions with the same mobility as under reducing conditions (Fig. 3c), indicating that the association between collectrin and B0AT1 as well as other transporters occurs through non-covalent interactions. Thus, collectrin co-localizes and specifically associates with a subgroup of renal amino acid transporters.

a, Renal brush border membrane proteins (100 μg) from collectrin+/y and collectrin-/y mice were incubated overnight with anti-B0AT1, anti-XT2, and anti-XT3 and anti-b0,+AT antibodies. Immunoprecipitated (IP) complexes were analysed by western blot using an anti-collectrin antibody (arrow). b, Collectrin co-localizes with B0AT1 in kidney proximal tubules. Kidney sections from wild-type mice were stained with antibodies to collectrin (green) and B0AT1 (red). Yellow indicates co-localization. S1, initial part of the proximal tubule. G, glomerulus. Magnifications, ×400. c, Western blot analysis of isolated renal brush border membranes under reducing (+DTT; 100 mM) and non-reducing (-DTT) conditions from collectrin+/y and collectrin-/y mice using antibodies directed against collectrin and B0AT1. Under reducing as well as under non-reducing conditions collectrin and B0AT1 appear as bands of ∼44 and ∼60 kDa, respectively (arrows). Molecular sizes are indicated in kDa.
To define the molecular mechanism of collectrin-regulated amino acid transport, we examined the effect of collectrin on B0AT1 function using Xenopus laevis oocytes and polarized MDCK cells. Two days of expression of B0AT1 alone in Xenopus oocytes conferred only a low transport activity for the B0AT1 substrate l-isoleucine16 (Fig. 4a). Collectrin alone did not exhibit any transport activity. However, co-expression of B0AT1 with collectrin resulted in a striking increase in l-isoleucine uptake (Fig. 4a). As a control, co-expression of collectrin with the Na+/phosphate IIa co-transporter in oocytes did not affect the rates and kinetics of phosphate uptake (Fig. 4b). To assess whether collectrin changes binding of l-isoleucine to its transporter B0AT1, we measured the apparent affinity (Km) for l-isoleucine uptake (Fig. 4c). Co-transport of l-isoleucine with Na+ generated a saturable, reversible inward current with an apparent Km for l-isoleucine of 0.99 ± 0.17 mM when B0AT1 was expressed alone, and of 0.78 ± 0.10 mM when B0AT1 was co-expressed with collectrin. Thus, the collectrin-induced increase in l-isoleucine transport is not due to a change in apparent affinity, but due to an increase in the maximal transport rate of B0AT1, probably as a result of increased surface expression. Indeed, expression of collectrin allowed functional surface expression of B0AT1 and apical uptake of l-isoleucine in polarized kidney MDCK cells (Fig. 4d). Thus, collectrin enhances functional surface expression of the SLC6 family transporter B0AT1.

a, Xenopus laevis oocytes were injected with murine B0AT1, collectrin, or B0AT1 plus collectrin RNA. l-isoleucine transport was determined two days after injection. Each bar represents mean transport activity ± s.d. (n = 15 oocytes analysed). b, Xenopus laevis oocytes were injected with murine Na+/Pi IIa alone or Na+/Pi IIa plus collectrin; phosphate transport (mean ± s.d.) was then determined at the indicated time points. n = 7 oocytes per sample. c, The half-maximal uptake rate Km of l-isoleucine by B0AT1 at six days of expression in the absence or presence of collectrin. Km was determined using two electrode voltage clamps at -50 mV using 0–10 mM of l-isoleucine in the presence of 100 mM Na+. Data was normalized to 10 mM l-isoleucine for n = 11 oocytes per group. d, MDCK cells were transduced with murine B0AT1, collectrin, B0AT1 plus collectrin, or empty vector (pLHCX). Bars represent the mean uptake of 300 μM l-isoleucine in 3 min ± s.d. *P < 0.001 (Student’s t-test).
We conclude that genetic inactivation of collectrin in mice results in a major defect in renal amino acid reabsorption. Collectrin controls protein expression and function of apical amino acid transporters in the brush border membranes of proximal tubules. Mechanistically, collectrin associates with several amino acid transporters, enhances their surface expression, and thereby increases amino acid transport function. These data identify the ACE2 homologue collectrin as a regulator of renal amino acid uptake and may provide a molecular explanation for aminoaciduria in Hartnup disease, Fanconi syndrome, or diabetes.
Methods
collectrin knockout mice
A gene-targeting vector containing three loxP sites flanking exon 2 and a Neo selection cassette was constructed. Chimaeric progeny were crossed to the C57BL/6J Cre deleter line7 to generate complete null animals.
mRNA and protein analyses
Real-time polymerase chain reaction (PCR) was performed as described12. For protein analyses, total cell lysates and BBMVs of adult mouse kidney were prepared21 and probed with antibodies to mouse b0,+AT, NaPi IIa, aquaporin 2, collectrin, XT2, XT3, SIT1, B0AT1, ACE2, LAT2 and 4F2. For immunoprecipitations, brush border membrane proteins were probed with antibodies to B0AT1, XT2, XT3 and b0,+AT.
Urine and plasma analyses
Urine was collected over 24 h in a metabolic cage. Urinary crystals were observed under a magnification of ×1,000. For HPLC analysis19, crystals were dissolved in 0.1 M HCl. Na+, K+ and Cl- were assayed by indirect potentiometry. Phosphate was determined using the phosphomolybdate method. Calcium levels were obtained by cresolphthalein. Glucose, urea and uric acid were measured using Reflovet (Roche).
Histology and immunohistochemistry
Paraffin sections were stained with haematoxylin and eosin (H&E). Immunohistochemistry was performed with an affinity purified anti-collectrin antibody. B0AT1 was detected using a rabbit anti-mouse B0AT1 antibody.
Amino acid transporter activities
Uptake of 3H-Phe, 14C-Gln and 32PO4 was determined in BBMVs at 60 s. The final amino acid concentration was 1 mM. Measurements were done in parallel in the presence of either Na+ or K+. Expression studies and uptake measurements in Xenopus laevis oocytes and kidney MDCK cells were performed as described22,23.
References
Imai, Y. et al. Angiotensin-converting enzyme 2 protects from severe acute lung failure. Nature 436, 112–116 (2005)
Crackower, M. A. et al. Angiotensin-converting enzyme 2 is an essential regulator of heart function. Nature 417, 822–828 (2002)
Kuba, K. et al. A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus-induced lung injury. Nature Med. 11, 875–879 (2005)
Zhang, H. et al. Collectrin, a collecting duct-specific transmembrane glycoprotein, is a novel homolog of ACE2 and is developmentally regulated in embryonic kidneys. J. Biol. Chem. 276, 17132–17139 (2001)
Fukui, K. et al. The HNF-1 target Collectrin controls insulin exocytosis by SNARE complex formation. Cell Metabol. 2, 373–384 (2005)
Akpinar, P., Kuwajima, S., Krutzfeldt, J. & Stoffel, M. Tmem27: A cleaved and shed plasma membrane protein that stimulates pancreatic β cell proliferation. Cell Metabol. 2, 385–397 (2005)
Schwenk, F., Baron, U. & Rajewsky, K. A cre-transgenic mouse strain for the ubiquitous deletion of loxP-flanked gene segments including deletion in germ cells. Nucleic Acids Res. 23, 5080–5081 (1995)
Bergeron, A., Jorquera, R. & Tanguay, R. M. Hereditary tyrosinemia: an endoplasmic reticulum stress disorder?. Med. Sci. 10, 976–980 (2003)
Endo, F. et al. Animal models reveal pathophysiologies of tyrosinemias. J. Nutr. 3, 2063S–2067S (2003)
Verrey, F. et al. Novel renal amino acid transporters. Annu. Rev. Physiol. 67, 557–572 (2005)
Kenny, A. J. & Maroux, S. Topology of microvillar membrance hydrolases of kidney and intestine. Physiol. Rev. 62, 91–128 (1982)
Romeo, E. et al. Luminal kidney and intestine SLC6 amino acid transporters of B0AT-cluster and their tissue distribution in Mus musculus. Am. J. Physiol. Renal Physiol. 290, F376–F383 (2006)
Broer, A. et al. Molecular cloning of mouse amino acid transport system B0, a neutral amino acid transporter related to Hartnup disorder. J. Biol. Chem. 279, 24467–24476 (2004)
Kleta, R. et al. Mutations in SLC6A19, encoding B0AT1, cause Hartnup disorder. Nature Genet. 6, 999–1002 (2004)
Seow, H. F. et al. Hartnup disorder is caused by mutations in the gene encoding the neutral amino acid transporter SLC6A19. Nature Genet. 6, 1003–1007 (2004)
Camargo, S. M., Makrides, V., Virkki, L. V., Forster, I. C. & Verrey, F. Steady-state kinetic characterization of the mouse BoAT1 sodium-dependent neutral amino acid transporter. Eur. J. Physiol. 451, 338–348 (2005)
Kowalczuk, S. et al. Molecular cloning of the mouse IMINO system: an Na+- and Cl--dependent proline transporter. Biochem. J. Mar. 15, 417–422 (2005)
Peghini, P., Janzen, J. & Stoffel, W. Glutamate transporter EAAC-1-deficient mice develop dicarboxylic aminoaciduria and behavioral abnormalities but no neurodegeneration. EMBO J. 16, 3822–3832 (1997)
Feliubadalo, L. et al. Slc7a9-deficient mice develop cystinuria non-I and cystine urolithiasis. Hum. Mol. Genet. 12, 2097–2108 (2003)
Wagner, C. A., Lang, F. & Broer, S. Function and structure of heterodimeric amino acid transporters. Am. J. Physiol. Cell Physiol. 81, C1077–C1093 (2001)
Malathi, P., Preiser, H., Fairclough, P., Mallett, P. & Crane, R. K. A rapid method for the isolation of kidney brush border membranes. Biochim. Biophys. Acta 554, 259–263 (1979)
Ristic, Z. et al. Neutral amino acid transport mediated by ortholog of imino acid transporter SIT1/SLC6A20 in opossum kidney cells. Am. J. Physiol. Renal Physiol. 290, F880–F887 (2006)
Bauch, C., Forster, N., Loffing-Cueni, D., Summa, V. & Verrey, F. Functional cooperation of epithelial heteromeric amino acid transporters expressed in Madin–Darby canine kidney cells. J. Biol. Chem. 278, 1316–1322 (2003)
Acknowledgements
We thank M. J. Crackower, A. Leibbrandt, Y. Imai, K. Kuba and many others for their contributions. Supported by grants from The National Bank of Austria, The Austrian Ministry of Science and Education, IMBA, an EU Marie Curie Excellence grant, and EUGeneHeart to J.M.P., from the Swiss National Science Foundation to F.V. and EUGINDAT to F.V. and C.A.W.
Author Contributions U.D, R.S., C.R., C.B., G.S., A.R., S.A., J.A.P., D.S., S.M.R.C., V.M., T.R. and C.A.W. performed experiments. U.D., F.V., C.A.W. and J.M.P. planned experiments and wrote the manuscript.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
Reprints and permissions information is available at www.nature.com/reprints. The authors declare no competing financial interests.
Supplementary information
Supplementary information
This file contains Supplementary Methods, Supplementary Table and Supplementary Figures 1-7.
Rights and permissions
About this article
Cite this article
Danilczyk, U., Sarao, R., Remy, C. et al. Essential role for collectrin in renal amino acid transport. Nature 444, 1088–1091 (2006). https://doi.org/10.1038/nature05475
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature05475
This article is cited by
-
Amino acid sensor GCN2 promotes SARS-CoV-2 receptor ACE2 expression in response to amino acid deprivation
Communications Biology (2022)
-
The Lepidopteran KAAT1 and CAATCH1: Orthologs to Understand Structure–Function Relationships in Mammalian SLC6 Transporters
Neurochemical Research (2022)
-
ACE2 Nascence, trafficking, and SARS-CoV-2 pathogenesis: the saga continues
Human Genomics (2021)
-
Angiotensin converting enzyme 2 is a novel target of the γ-secretase complex
Scientific Reports (2021)
-
COVID-19-induced acute renal tubular injury associated with elevation of serum inflammatory cytokine
Clinical and Experimental Nephrology (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
