
Abstract
Arising from: X. Xu et al. Nature 421, 335–340 (2003); F. Zhang & Z. Zhou Nature 431, 925 (2004); X. Xu et al. reply; F. Zhang et al. reply
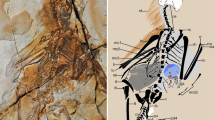
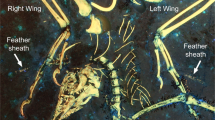
Our understanding of the origin of birds, feathers and flight has been greatly advanced by new discoveries of feathered non-avian dinosaurs, but functional analyses have not kept pace with taxonomic descriptions. Zhang and Zhou describe feathers on the tibiotarsus of a new basal enantiornithine bird from the Early Cretaceous of China1. They infer, as did Xu and colleagues from similar feathers on the small non-avian theropod Microraptor found in similar deposits2, that these leg feathers had aerodynamic properties and so might have been used in some kind of flight.
Similar content being viewed by others

Main
Claims that these were ‘four-winged’ dinosaurs that took to the air require investigations using independent lines of evidence, which was done by neither team of authors. There is no evidence to confirm the existence of a four-winged stage that involved feathers on the hindlimbs and that could have been an intermediate between feathered, ground-dwelling theropods and aerial birds.
First, the long leg feathers in question are said to preserve either asymmetrical vanes2 or little asymmetry but curved shafts1, from which the authors infer that these feathers had a residual aerodynamic function1. Neither feature indicates a function in lift or thrust (and therefore in flight). It has been suggested3 that both features are functionally correlated to flight, but the connection is not causal. Asymmetrical feathers resist torsion, and both features slightly reduce drag, which would be useful to a running animal with long feathers but would make no difference at realistic running speeds. Small non-avian theropods were good runners, but there is no evidence as yet that they flew.
Second, no evidence1,2 of how the feathers were attached to the hindlimb has been documented, so there is no indication that the feathers could have supported an aerodynamic load.
Third, Xu et al.2 offer no support for their statement that the hindlimbs (and therefore the inferred ‘hindwing’) of Microraptor extended laterally like those of bats, but unlike those of any bird or other theropod dinosaur. Furthermore, no explanation is given of how the leg could have been raised and extended into the lateral plane without dislocating the hip joints. Their remarkable assertion that the tibia of Microraptor is ‘bowed’ is also unsupported, and this is more likely to be the result of being crushed and of crossing other preserved bones. Even a revised posture of the hindlimbs does not mitigate the case for ‘four-winged gliding’ — it would also have to be subjected to functional tests.
Fourth, the proposed function of leg feathers in flight is unknown in birds of today, and the necessary structural modifications are not present in these extinct animals or in their relatives; therefore, the inferences drawn by the authors have no support from either the extinct or extant phylogenetic brackets4. Because leg feathers first appeared in non-flying theropods2, some of considerable size, their original function was not in flight, and no flight-related explanation for their presence in basal birds is necessary5. Long leg feathers are found in owls and grouse, and in nearly all raptors6: as with the feathers in these extinct forms, these are long, vaned and nearly symmetrical, but they have no demonstrated role in flight.
Fifth, the analogy that the authors draw1 between the use of webbed feet in kittiwakes as airbrakes and the extension of the hindlimbs in rapidly descending vultures1 is inappropriate: the first example induces drag, and the second relocates the centre of mass as the bird increases the angle of attack of the wings. Neither function uses or requires leg feathers, or produces lift. To be useful in flight, the advantage to lift must outweigh the disadvantage of drag.
Sixth, the authors should have calculated the mass, wing area, centre of mass and the centre of lift that would have been necessary to test whether the reconstruction of Microraptor was likely to be able to glide2. Such basic calculations must precede any assertion about gliding ability.
Most important, none of the evidence reported by the authors1,2 should have been advanced as support for an ‘arboreal’ origin of bird flight. It is recognized that the arboreal versus cursorial dichotomy of models for the origin of bird flight is not capable of resolution and should have been abandoned long ago7,8. Rather, the origin of the flight stroke is the central problem in the origin of flight, and so far nothing has been brought to light to indicate that Microraptor has any bearing on this question. Functional inferences should be based on functional studies, not just on analogies9. Ideas about arboreality should hinge on more than toe and claw proportions, and ideas about gliding on more than the profile of leg feathers.
References
Zhang, F. & Zhou, Z. Nature 431, 925 (2004).
Xu, X. et al. Nature 421, 335–340 (2003).
Feduccia, A. & Tordoff, H. B. Science 203, 1021–1022 (1979).
Witmer, L. D. in Functional Morphology in Vertebrate Paleontology (ed. Thomason, J. J.) 19–33 (Cambridge Univ. Press, 1995).
Gould, S. J. & Vrba, E. S. Paleobiology 8, 4–15 (1982).
Perrins, C. M. The Illustrated Encyclopedia of Birds (Prentice Hall, New York, 1990).
Padian, K. in New Perspectives on the Origin and Early Evolution of Birds (eds Gauthier, J. A. & Gall, L. F.) 255–272 (Yale Univ. Press, New Haven, 2001).
Dial, K. P. Science 299, 402–404 (2003).
Padian, K. Am. Zool. 41, 598–607 (2001).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Padian, K., Dial, K. Could ‘four-winged’ dinosaurs fly?. Nature 438, E3 (2005). https://doi.org/10.1038/nature04354
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature04354
This article is cited by
-
Dinosaur up in the air
Nature (2015)
-
IgH loci of American alligator and saltwater crocodile shed light on IgA evolution
Immunogenetics (2013)
-
The evolutionary continuum of limb function from early theropods to birds
Naturwissenschaften (2009)
-
A fundamental avian wing-stroke provides a new perspective on the evolution of flight
Nature (2008)
-
Could ‘four-winged’ dinosaurs fly? (Reply)
Nature (2005)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
