
Abstract
Understanding patterns of biodiversity distribution is essential to conservation strategies1, but severe data constraints make surrogate measures necessary2,3,4. For this reason, many studies have tested the performance of terrestrial vertebrates as surrogates for overall species diversity, but these tests have typically been limited to a single taxon or region3,4,5,6,7,8,9,10. Here we show that global patterns of richness are highly correlated among amphibians, reptiles, birds and mammals, as are endemism patterns. Furthermore, we demonstrate that although the correlation between global richness and endemism is low, aggregate regions selected for high levels of endemism capture significantly more species than expected by chance. Although areas high in endemism have long been targeted for the protection of narrow-ranging species11,12, our findings provide evidence that endemism is also a useful surrogate for the conservation of all terrestrial vertebrates.
Similar content being viewed by others


Main
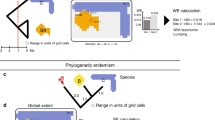
One of the challenges to preserving the Earth's biota is that species are unevenly distributed1. Confronted with the continuing extinction crisis, conservation strategies often focus either on areas with high species richness to maximize the number of species covered, or on areas that contain large numbers of endemic species (species found nowhere else)11,12,13,14 (Fig. 1a, b). The vast majority of species, however, have yet to be named, and information regarding their ranges (as well as the geographic ranges of many described species) is lacking15,16. Vertebrates, being relatively well-known, are frequently used to represent all biodiversity, but analyses of cross-taxa congruence often show little overlap5,6,7 and have thus lowered confidence in the use of surrogates. This lack of concordance could be a consequence of limitations in the taxonomic breadth or geographic extent of previous studies3,4,5,6,7,8,9,10.

These proportional indices combine the four terrestrial vertebrate classes and adjust for ecoregion area. Each scale bar of five colours represents relative levels of diversity from low (light) to high (dark). a, Species richness. b, Species endemism.
Here we use a uniquely comprehensive data set of terrestrial vertebrate distributions to evaluate global concordance in diversity patterns among four classes: amphibians, reptiles, birds and mammals. We recorded the presence or absence of 26,452 species according to 799 terrestrial ecoregions of the world17 (data available at http://www.worldwildlife.org/wildfinder). These ecoregions serve as the basis of World Wildlife Fund's conservation planning14, The Nature Conservancy's international efforts18, and the delineation of Conservation International's hotspots19 and high biodiversity wilderness areas20. Because ecoregions are created with species assemblages in mind, they are more useful units for comparing species data than are national lists21.
Richness is perhaps the most common measure of species diversity because it is relatively easy to compile and it implies that large numbers of species can be conserved in small areas1,9,13. We calculated the correlation between the proportional richness corrected for ecoregion area (hereafter termed ‘richness’; see Methods) of each class and the other vertebrates (an index combining the three remaining classes). Correlations between richness within a class and the richness of the other classes were strong, positive and significant for amphibians, birds and mammals (Pearson correlation coefficients, which are used throughout, are 0.591, 0.715 and 0.668, respectively; P < 0.01) (Table 1). A positive and significant correlation between reptiles and other vertebrates also existed, but was more moderate (0.380, P < 0.01) (Table 1). Ongoing debates over the causes of coincident patterns of global biodiversity include hypotheses regarding climate, geologic history, and a statistical consequence of differences in range size among species. All have explanatory merit, and a combination of factors seems likely22. Regardless of the underlying mechanisms, our results confirm that patterns of species richness among terrestrial vertebrates are broadly concordant.
Endemic species are another important target of global conservation efforts11,12. These species, often having small populations and few sites for conservation intervention, are inherently vulnerable to extinction23. As for species richness, a smaller amount of total area will need to be conserved if endemism patterns among taxa are correlated. We found that proportional endemism (hereafter termed ‘endemism’; see Methods) within each vertebrate class showed significantly positive correlations with endemism of the remaining three classes (Table 1). Endemism of reptiles (0.587, P < 0.01) and birds (0.612, P < 0.01) were more strongly related to their respective three-class index than endemism of mammals (0.490, P < 0.01) and amphibians (0.503, P < 0.01). These findings indicate that endemism within a well-documented group may be useful for guiding conservation decisions regarding overall endemism. Of course, correlations among classes only explain a portion of the observed variance in endemism among ecoregions. Therefore, conservation strategies based on one taxon will have to be supplemented with specific information for other groups in order to capture all endemic species.
Studies of congruence between richness and endemism have been inconclusive at regional scales8,24, and the one previous global test showed low congruence within birds8. Our analysis found no meaningful correlation between richness and endemism within any of the four terrestrial vertebrate classes (correlation coefficients -0.099 to 0.096) or for vertebrates overall (- 0.025) (Table 1), confirming that global patterns of these diversity measures are not spatially concordant. Thus, global conservation priorities based on richness alone will overlook many endemic species.
Although global correlations are suggestive of concordant diversity patterns, the question most relevant to conservation decisions is whether a specific set of ecoregions selected for one measure will represent non-target species3,4,25. Because endemic species are particularly important, we test how efficiently ecoregions chosen based on the level of endemism capture all species (Fig. 2a). After only 10% of the terrestrial land area is selected (an arbitrary cutoff4), the aggregated set of ecoregions contain 56.5% of the world's terrestrial vertebrate endemics. Of more interest here, these ecoregions capture 61.6% of all vertebrates, significantly higher than expected through random ecoregion selection (1,000 randomizations, 47.5 ± 2.2%, P < 0.01) (Supplementary Table 1 and Supplementary Fig. 1). The numbers of both endemics and all species captured continue to rise more steeply than expected, reaching an asymptote at roughly 50% of the Earth's surface (Fig. 2a). However, the actual area is much smaller than indicated by the analysis, simply because no ecoregion is pristine and most have large areas that are now of limited use for biodiversity. Selecting ecoregions on the basis of the endemism of a single class sometimes performs even better than using all four. For example, 10% of the world's land area chosen on the basis of bird endemism alone captures 59.6% of all vertebrate endemics; and on the basis of amphibian and reptile endemism, 71.0% and 72.0%, respectively, of total vertebrate species are captured (Supplementary Table 2). Our findings demonstrate that, although broad correlations between richness and endemism are weak, priority sets based on endemic species contain large numbers of total species. This result is probably due to turnover in species composition among areas of high endemism (that is, these areas are highly complementary in terms of non-endemic species26) and suggests that endemism is particularly useful for conservation prioritization.

a, The percentage of endemic species (red line) and all species (dark blue line) rise rapidly as the per cent of ecoregion area selected (blue shading) increases. The green line represents the total per cent of species captured when ecoregions are selected at random (mean of 1,000 random sets). Species accumulate significantly faster than expected by chance (lying above a 95% confidence interval, dashed line), and nearly all terrestrial vertebrates are found within ecoregions that represent 50% of the Earth's surface. b, Ecoregions representing 10% of the Earth's terrestrial surface (dark brown) that capture 56.5% of vertebrate endemics and 61.6% of all species. Also depicted are the additional ecoregions required to represent 50% of the Earth (orange); together, the two sets of ecoregions contain nearly all species of terrestrial vertebrates.
Although our work furthers the understanding of how species diversity patterns can inform conservation priorities at a global scale, we must point to several important caveats. The distribution patterns we report only apply to vertebrates and might not hold for more species-rich taxa such as plants, invertebrates and fungi. Biodiversity of vertebrates also encompasses aspects of population and genetic diversity that are missed when using species as the sole unit of measurement. Furthermore, methods for setting conservation priorities are complex and should consider not just the number of endemics or total species present, but also degree of threat7,11, population viability18, ecological and evolutionary processes14,18, and economic costs and benefits of conservation27. Comprehensive conservation strategies will require efforts at multiple scales to ensure the long-term survival of biodiversity in a region2,18. Using endemism along with other factors to identify global priorities helps to focus these conservation efforts on critical regions11,12,14, where on-the-ground efforts will yield the greatest payoffs for biodiversity.
Methods
Database
The database used for these analyses contains presence/absence data for the world's terrestrial amphibians (n = 4,797), reptiles (n = 7,483), birds (n = 9,470) and mammals (n = 4,702) by ecoregion. These data are based on the natural, historic ranges of extant species, such that species introduced, present as human commensals, vagrants or passage migrants were not recorded.
The terrestrial ecoregions are those delineated in ref. 17. Mangrove ecoregions were excluded from analysis because the land area of these regions is invariably small; hence species lists compiled from overlaying range maps inflates species totals. A further nine ecoregions were excluded from the analysis because of lack of data, but we are confident that these omissions do not affect overall distribution patterns (land area of the world's ecoregions is 134,735,751 km2, land area of the nine excluded ecoregions is 236,100 km2 or 0.175%). Because the large interior portions of Greenland and Antarctica contain no terrestrial vertebrates and are not mapped as ecoregions17, they too were excluded from the analysis as well as from calculations of Earth's land area.
Analyses
Class richness was tallied for each ecoregion and divided by the total number of species in the database for that class10 (or the total number within biomes when the data were regressed separately; see below). This proportional species richness allowed calculations to be comparable between taxa without a single species group overwhelming the others:

where e is the richness index for each ecoregion, n is the number of species groups used in the index, Gi(e) is the number of species in group i per ecoregion, and Gi(t) is the total number of species of group i. Endemism indices were calculated in the same manner. An adjusted richness index was used for comparisons between overall richness and endemism, whereby the richness totals included only non-endemic species so that endemics were not part of both comparative sets24.
We regressed each index against area (both variables were log10-transformed) in order to reduce the influence of ecoregion size on the indices. Because the relationship between species richness and area varied by biome, we performed regressions for each biome separately, and used the combined residuals of each index for subsequent analyses. There was, however, no significant relationship between the endemism indices and area, and we therefore used the original indices for endemism. There was also no effect of latitude after we accounted for area.
Pearson correlations were used for all analyses. We tested for statistical significance using a geographically restricted randomization approach with 10,000 iterations. Randomizations were constrained by both biome and biogeographic realm to reduce Type I errors owing to spatial autocorrelation28. This method is more stringent than unrestricted randomizations or randomizations restricted to either biomes or realms28. We used the following standards to interpret correlation coefficients: a large correlation coefficient was approximately 0.50 and higher, moderate correlations were around 0.30, and small correlations were about 0.10 (refs 29, 30).
To determine whether the percentage of species captured when selecting ecoregions for endemism was greater than by chance alone, we selected 1,000 random sets of ecoregions for every 10% interval in area, up to 67.9% (the percentage at which all species were captured). Each iteration selected the same number of ecoregions as in the set chosen for endemism at the corresponding interval (see Supplementary Information). Note that because ecoregions vary in size, the percentages of Earth's land surface are slightly less than the 10% breaks. For example, ecoregions selected for the most endemics within 10% of the global land area actually make up 9.29% (always favouring the more conservative number).
References
Gaston, K. J. Global patterns in biodiversity. Nature 405, 220–227 (2000)
Margules, C. R. & Pressey, R. L. Systematic conservation planning. Nature 405, 243–253 (2000)
Howard, P. C. et al. Complementarity and the use of indicator groups for reserve selection in Uganda. Nature 394, 472–475 (1998)
Moore, J. L. et al. Performance of sub-Saharan vertebrates as indicator groups for identifying priority areas for conservation. Conserv. Biol. 17, 207–218 (2003)
Prendergast, J. R., Quinn, R. M., Lawton, J. H., Eversham, B. C. & Gibbons, D. W. Rare species, the coincidence of diversity hotspots and conservation strategies. Nature 365, 335–337 (1993)
van Jaarsveld, A. S. et al. Biodiversity assessment and conservation strategies. Science 279, 2106–2108 (1998)
Dobson, A. P., Rodriguez, J. P., Roberts, W. M. & Wilcove, D. S. Geographic distribution of endangered species in the United States. Science 275, 550–553 (1997)
Orme, C. D. L. et al. Global hotspots of species richness are not congruent with endemism or threat. Nature 436, 1016–1019 (2005)
Brooks, T. et al. Toward a blueprint for conservation in Africa. Bioscience 51, 613–624 (2001)
Ricketts, T. H., Dinerstein, E., Olson, D. M. & Loucks, C. Who's where in North America? Bioscience 49, 369–381 (1999)
Myers, N., Mittermeier, R. A., Mittermeier, C. G., da Fonseca, G. A. B. & Kent, J. Biodiversity hotspots for conservation priorities. Nature 403, 853–858 (2000)
Stattersfield, A. J., Crosby, M. J., Long, A. J. & Wege, D. C. Endemic Bird Areas of the World: Priorities for Biodiversity Conservation (BirdLife International, Cambridge, 1998)
Mittermeier, R. A., Robles Gil, P. & Mittermeier, C. G. Megadiversity: Earth's Biologically Wealthiest Nations (CEMEX, Mexico City, 1997)
Olson, D. M. & Dinerstein, E. The global 200: A representation approach to conserving the Earth's most biologically valuable ecoregions. Conserv. Biol. 12, 502–515 (1998)
Raven, P. H. & Wilson, E. O. A fifty-year plan for biodiversity surveys. Science 258, 1099–1100 (1992)
Millennium Ecosystem Assessment. Ecosystems and Human Well-being: Biodiversity Synthesis (World Resources Institute, Washington DC, 2005)
Olson, D. M. et al. Terrestrial ecoregions of the world: A new map of life on Earth. Bioscience 51, 933–938 (2001)
Groves, C. R. Drafting a Conservation Blueprint: A Practitioner's Guide to Planning for Biodiversity (Island Press, Washington DC, 2003)
Mittermeier, R. A. et al. Hotspots Revisited (CEMEX, Mexico City, 2004)
Mittermeier, R. A. et al. Wilderness and biodiversity conservation. Proc. Natl Acad. Sci. USA 100, 10309–10313 (2003)
Pimm, S. L. Seeing both the woods and the trees. Nature 402, 853–854 (1999)
Pimm, S. L. & Brown, J. H. Domains of diversity. Science 304, 831–833 (2004)
Gaston, K. J. Rarity as double jeopardy. Nature 394, 229–230 (1998)
Ricketts, T. H. Aligning conservation goals: are patterns of species richness and endemism concordant at regional scales? Anim. Biodivers. Conserv. 24, 91–99 (2001)
Balmford, A. On hotspots and the use of indicators for reserve selection. Trends Ecol. Evol. 13, 409 (1998)
Williams, P. H. in Conservation in a Changing World (eds Mace, G. M., Balmford, A. & Ginsberg, J. R.) 211–249 (Cambridge Univ. Press, Cambridge, 1998)
Balmford, A. et al. Economic reasons for conserving wild nature. Science 297, 950–953 (2002)
Manly, B. F. J. Randomization, Bootstrap and Monte Carlo Methods in Biology 2nd edn (Chapman and Hall, London, 1997)
Aron, A. & Aron, E. N. Statistics for Psychology 3rd edn (Prentice Hall, Upper Saddle River, New Jersey, 2003)
Cohen, J. Statistical Power Analysis for the Behavioral Sciences (Erlbaum, Hillsdale, New Jersey, 1988)
Acknowledgements
This work was funded by World Wildlife Fund's Living Planet Campaign with support from J. Leape. We thank P. Ehrlich, K. Gaston and H. Possingham for comments; and the following for expertise, unpublished data and technical assistance: A. Allison, T. Allnutt, S. Anderson, L. Aquino, P. Beresford, G. Brennan, D. Broadley, R. Brown, C. Burdette, N. Burgess, L. Burrus, J. Campbell, F. Castro, L. Cayet, E. Colijn, L. Coloma, R. Crombie, I. Das, I. de La Riva, A. Diesmos, W. Duellman, K. Ernst, M. Evans, L. Fitzgerald, J. Fjeldså, J. Formas, D. Frost, S. Galster, A. Gascoigne, T. Grant, M. Hadjichristoforou, J. D'Amico Hales, L. Hansen, R. Heyer, R. Hoffmann, M. Hoogmoed, R. Ibanez, J. Icochea, D. Iskandar, N. Jorge da Silva, G. Köhler, C. Kurtonur, E. La Marca, J. Langone, E. Lavilla, F. Lobo, S. Lötters, C. Loucks, S. Madge, O. Marques, M. Martins, J. McCranie, R. McDiarmid, R. Montero, J. Ortiz, S. Palminteri, J. Patterson, J. Pilgrim, G. Pinto, J. Porter, G. Powell, C. Rahbek, S. Reichle, E. Richard, G. Rodda, J. Savage, E. Schaub, J. Schipper, N. Scott, G. Scrocchi, W. Sechrest, H. Strand, S. Stuart, M. Taye, R. Tizard, D. Turney, D. Urban, P. Uetz, E. Underwood, P. van Dijk, O. Villela, J. Weiss, W. Wettengel, L. Wilson, Y. Xie, A. Yanosky, E. Zhao and G. Zug. We also thank the Species Survival Commission of IUCN, The Nature Conservancy and the Zoological Museum of the University of Copenhagen for data contributions.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
Reprints and permissions information is available at npg.nature.com/reprintsandpermissions. The authors declare no competing financial interests.
Supplementary information
Supplementary Notes
This file contains Supplementary Figure 1, Supplementary Methods and Supplementary Discussions on unadjusted richness and endemism, and ecoregions selected on the basis of endemism versus random procedures. (DOC 60 kb)
Supplementary Table 1
This table lists the percentage of endemic species and total species captured at each ten percent interval of Earth’s land area up to 67.9% when selecting ecoregions for endemism and by random. The percent of both endemics and of all species captured is higher when selecting on the basis of endemism. (DOC 34 kb)
Supplementary Table 2
This table lists the percentage of endemic and total species captured when selecting ecoregions for the highest levels of endemism in each class until 10% of the Earth’s land area was reached. All four selections included more than half of the species contained in the three remaining groups, and all but mammals contained about half of the endemics. These findings emphasize the point that the areas of greatest conservation concern for any one species group are crucial for other species. (DOC 29 kb)
Rights and permissions
About this article
Cite this article
Lamoreux, J., Morrison, J., Ricketts, T. et al. Global tests of biodiversity concordance and the importance of endemism. Nature 440, 212–214 (2006). https://doi.org/10.1038/nature04291
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature04291
This article is cited by
-
There is a trade-off between forest productivity and animal biodiversity in Europe
Biodiversity and Conservation (2023)
-
M3=Maths on Morphometry of Moths
Resonance (2023)
-
Relocating croplands could drastically reduce the environmental impacts of global food production
Communications Earth & Environment (2022)
-
Preliminary survey of non-human Primates in four forest areas of Yewa River Basin at the Nigeria-Benin Republic transboundary
Biologia (2022)
-
Population size and habitat preference of the Omo River guereza (Colobus guereza guereza) in a multi-habitat matrix in the central highlands of Ethiopia
Primates (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
