
Abstract
Arising from: J. Mecsas et al. Nature 427, 606 (2004) Mecsas and colleagues suggest that a deficiency in the chemokine receptor CCR5 in humans is unlikely to confer protection against plague, based on their study of Yersinia pestis infection in Ccr5-deficient mice1. They were testing the hypothesis that a mutation in the CCR5 gene, frequently found in Caucasians, may have been selected for in the past because it provided protection against (bubonic) plague2,3,4,5,6,7; the mutation, called CCR5Δ32, is characterized by a 32-base-pair deletion. We have also tested this hypothesis by using Y. pestis infection in mice and, in addition, we have done phagocytosis experiments with macrophages from wild-type and Ccr5-deficient mice. Although, like Mecsas et al., we did not see any difference in the survival of the two groups of mice, we did find that there was a significantly reduced uptake of Y. pestis by Ccr5-deficient macrophages in vitro. Our results indicate that the role of Ccr5 in Y. pestis infection may therefore be more complex than previously thought.
Similar content being viewed by others

Main
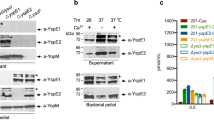
In humans, macrophages are targeted by Y. pestis, the causative agent of plague, and are therefore important for successful infection. We tested whether Ccr5 affects the uptake of Y. pestis by macrophages in vitro by using peritoneal macrophages from Ccr5+/+ and Ccr5−/− mice in phagocytosis assays. The uptake by Ccr5−/− macrophages was about 30-fold lower than that by Ccr5+/+ macrophages (Fig. 1a; six independent experiments). Our preliminary results indicate that the uptake of Yersinia pseudotuberculosis by macrophages from Ccr5−/− mice is much less inhibited in similar experiments (Fig. 1b), suggesting that the inhibition may be specific to Y. pestis.

a, b, The intracellular bacteria recovered from peritoneal macrophages isolated from C57BL/6 Ccr5+/+ and Ccr5−/− mice and incubated (1×106 cells for 1 h at 37°C) with a, Y. pestis GB (multiplicity of infection, 10 colony-forming units (CFU); mean±s.e.m.) or b, Y. pseudotuberculosis strain IP32953. Gentamycin was used to kill extracellular bacteria. c, Survival of C57BL/6 Ccr5+/+ mice (blue; n = 9) and Ccr5−/− mice (orange; n = 10) after challenge with 2 CFU Y. pestis GB (Biovar orientalis, Pgm+, LcrV+; median lethal dose is 1 CFU) subcutaneously.
To test the effect of Ccr5 on survival after Y. pestis infection, groups of specific pathogen-free Ccr5+/+ and Ccr5−/− mice were challenged with lethal inocula of Y. pestis GB, a highly virulent strain isolated from a fatal human case of plague. However, there was no significant difference in survival between the groups, even after infection with a low dose of two colony-forming units (CFU) (Fig. 1c).
Our survival data are in agreement with those of Mecsas et al.1, although we used a strain of Y. pestis with a different degree of virulence (GB rather than KIM), mice with a different genetic background (C57BL/6 rather than BALB/c) and a different route of infection (subcutaneous rather than intravenous). Our results show that Ccr5−/− mice are not protected against infection with a fatal human isolate of Y. pestis and succumb at the same rate as Ccr5+/+ mice.
Although these results seem to disprove the ‘plague hypothesis’, some doubts remain. We consistently observed a marked reduction in the uptake of Y. pestis by Ccr5−/− macrophages in vitro that appears to be specific to this species of Yersinia. The Y. pestis strain that caused the great plague pandemic in the fourteenth century was probably quite different from the twentieth-century isolate used for the infection experiments discussed here. Genome analysis indicates that Y. pestis evolved rapidly from an enteric organism, which was spread by the faecal–oral route, to a flea-transmitted pathogen of rodents and humans, with acquisition of novel virulence mechanisms along the way8,9.
In addition, the pathogenesis of Y. pestis infection may not be comparable when delivered by injection of mice in the laboratory rather than by flea-borne transmission to humans10, because infection may be more rapid and acute. The dose of plague bacteria delivered by flea-borne transmission is likely to be more variable and the outcome of infection to depend on an interaction between the pathogen, vector and mammalian host. A previous infection leading to preactivation of the host's immune system would change the course of a subsequent Y. pestis infection — as would be expected in people living in the Middle Ages, who were constantly encountering all kinds of infection and in whom a resistance to plague could have developed in association with the CCR5Δ32 mutation.
Under these circumstances, firm conclusions cannot be drawn from the negative results obtained in Ccr5-deficient mice. Taking all these arguments into consideration, the data on the role of CCR5 in Y. pestis infection are still inconclusive because the situation seems to be more complex than previously anticipated.
References
Mecsas, J. et al. Nature 427, 606 (2004).
Blanpain, C., Libert, F., Vassart, G. & Parmentier, M. Recept. Chann. 8, 19–31 (2002).
Samson, M. et al. Nature 382, 722–725 (1996).
Liu, R. et al. Cell 86, 367–377 (1996).
Dean, M. et al. Science 273, 1856–1862 (1996).
Stephens, J. C. et al. Am. J. Hum. Genet. 62, 1507–1515 (1998).
Libert, F. et al. Hum. Mol. Genet. 7, 399–406 (1998).
Parkhill, J. et al. Nature 413, 523–527 (2001).
Wren, B. W. Nature Rev. Microbiol. 1, 55–64 (2003).
Jarrett, C. O., Sebbane, F., Adamovicz, J. J., Andrews, G. P. & Hinnebusch, B. J. Infect. Immun. 72, 2052–2056 (2004).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Elvin, S., Williamson, E., Scott, J. et al. Ambiguous role of CCR5 in Y. pestis infection. Nature 430, 418 (2004). https://doi.org/10.1038/nature02822
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature02822
This article is cited by
-
Plague as a cause for familial Mediterranean fever
Nature Immunology (2020)
-
FPR1 is the plague receptor on host immune cells
Nature (2019)
-
The many and varied niches occupied by Yersinia pestis as an arthropod-vectored zoonotic pathogen
Antonie van Leeuwenhoek (2005)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
