
Abstract
This study was designed to test the interaction between amyloid-β and tau proteins as a determinant of metabolic decline in preclinical Alzheimer’s disease (AD). We assessed 120 cognitively normal individuals with [18F]florbetapir positron emission tomography (PET) and cerebrospinal fluid (CSF) measurements at baseline, as well as [18F]fluorodeoxyglucose ([18F]FDG) PET at baseline and at 24 months. A voxel-based interaction model was built to test the associations between continuous measurements of CSF biomarkers, [18F]florbetapir and [18F]FDG standardized uptake value ratios (SUVR). We found that the synergistic interaction between [18F]florbetapir SUVR and CSF phosphorylated tau (p-tau) measurements, rather than the sum of their independent effects, was associated with a 24-month metabolic decline in basal and mesial temporal, orbitofrontal, and anterior and posterior cingulate cortices (P<0.001). In contrast, interactions using CSF amyloid-β1–42 and total tau biomarkers did not associate with metabolic decline over a time frame of 24 months. The interaction found in this study further support the framework that amyloid-β and hyperphosphorylated tau aggregates synergistically interact to cause downstream AD neurodegeneration. In fact, the regions displaying the metabolic decline reported here were confined to brain networks affected early by amyloid-β plaques and neurofibrillary tangles. Preventive clinical trials may benefit from using a combination of amyloid-β PET and p-tau biomarkers to enrich study populations of cognitively normal subjects with a high probability of disease progression in studies, using [18F]FDG as a biomarker of efficacy.
Similar content being viewed by others

Introduction
Alzheimer’s disease (AD) clinically develops in the context of presymptomatic, mild cognitive impairment and dementia stages, and can be quantified in vivo via imaging and fluid biomarkers.1 AD pathophysiological process has been modelled as a cascade of progressive neuropathological events initiated by amyloidosis followed by hyperphosphorylated tau accumulation and subsequent structural, functional and cognitive declines.1, 2 Recent longitudinal studies have shown that the coexistence of upstream events of the AD cascade predicts downstream structural and functional declines.3 For example, Knopman et al.4 have reported that faster structural and metabolic declines are observed in those individuals with abnormal baseline amyloid-β load and metabolism or hippocampal volume. Furthermore, studies conducted in cognitively normal individuals revealed amnestic and non amnestic cognitive declines in individuals with co-occurrence of abnormal amyloid-β load and cerebrospinal fluid (CSF) tau pathology, brain atrophy or hypometabolism.5, 6, 7, 8
A conceptual framework proposes that amyloid-β and tau pathologies synergistically potentiates subsequent downstream neurodegeneration.9 This synergistic framework challenges previous AD pathophysiological theories emphasising amyloid-β or tau pathologies as the major driving forces of disease progression, the so-called baptist and tauist perspectives, respectively.10 In fact, a cross-sectional study conducted in cognitive normal individuals suggested brain atrophy of temporoparietal and occipital cortices as a function of the interaction between abnormal CSF amyloid-β1–42 and phosphorylated tau (p-tau) biomarker status.11 Similarly, the deleterious associations between CSF amyloid-β1–42 and brain structure or cognition are dependent on the presence of abnormal CSF p-tau levels.12, 13 In fact, data supporting molecular synergistic interactions between amyloid-β and tau peptides, leading to downstream toxicity, have been described in the recent animal model literature.14, 15, 16
[18F]Fluorodeoxyglucose ([18F]FDG) positron emission tomography (PET) is one of the most important biomarkers in AD research and clinical practice. [18F]FDG abnormalities reflecting focal cerebral hypometabolism constitute a well-validated and sensitive biomarker associated with AD pathophysiology, with potential applications in following disease progression and assessing the efficacy of disease-modifying interventions.17 Although brain metabolism is vulnerable to AD pathophysiology, the relationship between amyloid-β and tau pathologies as determinants of brain hypometabolism is still unclear. Few longitudinal imaging studies have supported a link, albeit a modest one, between amyloid-β or tau pathology and metabolic decline.18, 19 Although the pathogenic synergism between brain amyloidosis and tau pathology has been postulated as a determinant of AD progression, no previous study so far has been tested whether the interactions between amyloid-β and tau pathologies are associated with developing brain hypometabolism.
Here, in a longitudinal study of cognitively normal elderly individuals, we test the hypothesis that [18F]FDG metabolic decline depends on the synergistic interaction between, rather than the sum of independent effects of, amyloid-β and tau biomarkers.
Materials and methods
Database description and study participants
Data used in the preparation of this article were obtained from the AD Neuroimaging Initiative (ADNI) database (adni.loni.usc.edu). The ADNI was launched in 2003 as a public–private partnership, led by Principal Investigator Michael W Weiner, MD. The primary goal of ADNI has been to test whether serial magnetic resonance imaging (MRI), PET, other biological markers, and clinical and neuropsychological assessment can be combined to measure the progression of mild cognitive impairment and early AD.
For the present study, we selected cognitively normal subjects who had undergone baseline lumbar puncture and [18F]florbetapir PET imaging, as well as both baseline and 24-month follow-up visits for the [18F]FDG PET. The operational definition of cognitively normal individuals adopted in this study was a mini-mental state examination score of 24 or higher, a clinical dementia rating 0 and absence of any neuropsychiatric diagnosis, including mild cognitive impairment and dementia. The inclusion/exclusion criteria adopted by the ADNI are described in detail at www.adni-info.org (accessed January 2016).
CSF analyses
CSF amyloid-β1–42, tau phosphorylated at threonine 181 and total tau were quantified using the multiplex xMAP Luminex platform (Luminex, Austin, TX, USA) with INNOBIA AlzBio3; (Innogenetics, Ghent, Belgium) immunoassay kit-based reagents. The CSF biomarker data sets used in this study were obtained from the ADNI files ‘UPENNBIOMK5-8.csv’, and all the biomarkers values for each individual were selected from the same file. Further details of ADNI methods for CSF acquisition and CSF measurement can be found at www.adni-info.org (accessed January 2016).
MRI/PET methods
ADNI, MRI and PET standard acquisition protocols are detailed elsewhere (http://adni.loni.usc.edu/methods; accessed January 2016). Imaging analysis methods are summarized in Figure 1. T1-weighted MRI images were corrected for field distortions and subsequently processed using the CIVET image-processing pipeline. In summary, images underwent non-uniformity correction, followed by brain masking and segmentation using the brain extraction based on nonlocal segmentation technique.20 Images were subsequently co-registered using a nine-parameter affine transformation and non-linearly spatially normalized to the MNI 152 reference template. PET images were blurred with a volumetric Gaussian kernel with a full-width half-maximum of 8 mm. Subsequently, linear co-registration and non-linear spatial normalization to the MNI 152 template space were performed using the transformation derived from the semiautomatic PET/T1-MRI transformation and anatomical MRI registration for each subject. Voxel-wise, standardized uptake value ratio (SUVR) maps were then generated for [18F]florbetapir using the cerebellum grey matter and the global white matter as reference regions. [18F]FDG SUVR images were generated using the pons as a reference region. A global SUVR value for each subject was estimated from the precuneus, as well as from the prefrontal, orbitofrontal, parietal, temporal, anterior and posterior cingulate cortices. Individual [18F]FDG SUVR values were obtained from the region of interest, tailored by voxel-based statistical clusters.

Summary of image analysis methods.
Statistical methods
Statistical analyses were performed using the R Statistical Software Package version 3.0.2 with the RMINC library (http://www.r-project.org/; accessed January 2016). RMINC is an imaging package that allows image files in the Medical Image NetCDF (MINC) format to be analysed within the R statistical environment.
The primary end point on this study was the rate of metabolic decline defined as:

The biomarkers were analyzed using z-scores. Simple linear regression models evaluated the effects of global [18F]florbetapir SUVR or CSF p-tau as single regressors on Δ%[18F]FDG. The voxel-based interaction model described below was built to test whether main and interactive effects between global [18F]florbetapir SUVR and CSF p-tau are associated with metabolic decline.

To ensure that our results were not due to issues related to voxel-based analysis, we then isolated the clusters, averaged the SUVRs and performed the same analysis for each cluster, using the averaged SUVRs as the outcomes. The model was adjusted for age, gender and APOE ɛ4 status.
The statistical parametric maps presented in this study were corrected for multiple testing. Statistical significance was defined using a false discovery rate with a threshold of P<0.001.
Results
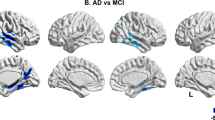
Demographics and key sample characteristics are summarized in Table 1. The effects of [18F]florbetapir SUVR or CSF p-tau as single regressors on metabolic decline were not significant in our population. Voxel-based analysis revealed that longitudinal [18F]FDG SUVR decline on basal and mesial temporal, orbitofrontal, anterior and posterior cingulate cortices was driven by the synergistic interaction between [18F]florbetapir SUVR and CSF p-tau measurements (Figure 2). Although the interaction term was significant (β3), the main effects of [18F]florbetapir SUVR (β1) and CSF p-tau (β2) on 24 months metabolic decline were not significant using our voxel-based regression model.

Synergistic effect between [18F]florbetapir SUVR and CSF p-tau drives [18F]FDG uptake decline in limbic regions. Statistical parametric maps, after correcting for multiple comparisons (false discovery rate corrected at P<0.001), overlaid in a structural MRI scan, reveal areas in which 24-month [18F]FDG uptake decline occurs as a function of the synergistic interaction between baseline [18F]florbetapir SUVR and CSF p-tau measurements. Significant interactive effects were observed in the basal and mesial temporal, orbitofrontal, and anterior and posterior cingulate cortices. The analysis was corrected for age, gender and APOE ɛ4 status. CSF, cerebrospinal fluid; [18F]FDG, [18F]fluorodeoxyglucose; MRI, magnetic resonance imaging; p-tau, phosphorylated tau; SUVR, standardized uptake value ratio.
Subsequently, we examined the interaction between [18F]florbetapir SUVR and CSF p-tau on Δ%[18F]FDG SUVR in the regions revealed by the voxel-based analysis, using an averaged SUVR value for each region of interest. The effect of the interaction on metabolic decline was higher in mesiobasal temporal (slope coefficient (β3)=−4.42, s.e.=1.16, P<0.0001) and orbitofrontal (slope coefficient (β3)=−4.12, s.e.=1.4, P=0.0002), followed by anterior (slope coefficient (β3)=−2.77, s.e.=0.73, P=0.0001) and posterior cingulate (slope coefficient (β3)=−1.78, s.e.=0.75, P=0.0009) clusters. The main effects of [18F]florbetapir SUVR and CSF p-tau on metabolic decline were not significant in each of the regions of interest evaluated. This interaction was absent in other brain regions such as the occipital lobe (slope coefficient (β3)=0.46, s.e.=1.03, P=0.65) and precuneus (slope coefficient (β3)=1.18, s.e.=0.73, P=0.14). Importantly, the interaction between [18F]florbetapir SUVR and CSF p-tau as a function of declines in [18F]FDG was not observed, using global [18F]FDG SUVR measurements (slope-coefficient (β3)=−0.51, s.e.=0.59, P=0.36).
Interestingly, the average global [18F]FDG SUVR decline in our population was 1.6% (95% confidence interval : 0.6–2.5%), whereas within the cluster revealed by the interaction analysis in basal and mesial temporal, orbitofrontal, anterior and posterior cingulate cortices the average [18F]FDG SUVR decline was 6.1% (95% confidence interval : 4.7–6.9%) over 24 months (Supplementary Figure 1).
In our study population, [18F]florbetapir SUVR was highly correlated with CSF amyloid-β1–42 (Spearman's rho=0.68, P<0.0001), whereas CSF p-tau was highly correlated with CSF total tau (Spearman’s rho=0.70, P<0.0001). However, alternative models showed that [18F]FDG SUVR decline was not associated with the interaction between CSF amyloid-β1–42 with total tau or p-tau. Furthermore, [18F]FDG SUVR decline was not associated with the interaction between CSF total tau and [18F]florbetapir SUVR. Similarly, interactions using CSF amyloid-β1–42 or total tau biomarkers were not significant in the mesiobasal temporal, orbitofrontal, anterior and posterior cingulate clusters.
Notably, longitudinal brain hypometabolism was independent of APOE ɛ4 status or its interactions with CSF p-tau (slope coefficient (β3)=−0.47, s.e.=1.40, P=0.74), CSF total tau (slope coefficient (β3)=0.56, s.e.=1.37, P=0.69), CSF amyloid-β1–42 (slope coefficient (β3)=−0.70, s.e.=1.26, P=0.58) or [18F]florbetapir SUVR (slope coefficient (β3)=0.48, s.e.=1.31, P=0.72).
Discussion
The present study shows that the synergism between, rather than the sum of independent effects of, continuous brain amyloid-β deposition and p-tau biomarkers drives the rate of metabolic decline in AD-related regions in a cognitively normal elderly population. We further found that the interaction between brain amyloid-β deposition and total tau biomarkers did not predict metabolic decline.
The synergistic interaction between brain amyloid-β deposition and CSF p-tau biomarkers concentrations found in our study has to be carefully interpreted from the pathophysiological perspective. In the statistical model proposed, in terms of such interaction within the study population, the density of the brain amyloid-β deposition and CSF concentrations of p-tau synergistically amplify the severity of the subsequent brain hypometabolism. As such, individuals with the highest baseline concentrations of [18F]florbetapir SUVR and p-tau had the highest rate of metabolic decline in AD-related regions over a time frame of 24 months. One might claim that this interaction supports the conceptual framework that amyloid-β and hyperphosphorylated tau aggregates synergistically, rather than independently, potentiate downstream neurodegeneration.9, 21, 22 The combined effect between amyloid-β and hyperphosphorylated tau has been supported by animal model literature.23 For example, p-tau potentiates amyloid-β-induced neurotoxicity24 and molecular interactions between amyloid-β and p-tau have shown to amplify synaptic and neuronal damage in AD.25 Indeed, deleterious interactions between peptide aggregates have already been described in other proteinopathies such as Lewy body disease (alpha-synuclein)26 and frontal temporal lobar degeneration (TDP-43 inclusions).27
Fast metabolic decline in mesial temporal regions is not considered part of the normal aging process.28, 29 Indeed, the mesial temporal metabolic decline found in the present study is consistent with the idea that mesial temporal hypometabolism may be an early metabolic abnormality linked to the AD pathophysiological process.30, 31 Furthermore, the hypometabolic areas reported here converge to the brain circuits that are affected early by neurofibrillary tangles and amyloid-β plaques.32, 33, 34 Such convergence between metabolic and neuropathological abnormalities in limbic regions further supports a synergism between amyloid-β and tau pathologies leading to hypometabolism. Recently, a hypothetical model in which the interaction between amyloid-β and tau aggregates potentiates mesial temporal synaptic dysfunction in the preclinical stages of AD has been suggested.35 One may claim that brain amyloid-β measurements represent a proxy of oligomeric amyloid-β.36 As such, the deleterious effects of amyloidosis reported here might be mediated by amyloid-β oligomers rather than fibrillar conformations.
In addition to recent findings supporting the claim that the synergism between brain amyloidosis and neurodegeneration is associated with AD progression;3, 4 here we demonstrate that [18F]florbetapir PET rather than CSF amyloid-β1–42—and CSF p-tau rather than total tau—better predict mesial temporal metabolic decline. The motivation to explore these effects using amyloid-PET in our study is derived from its close association with Aβ plaques load,37 whereas CSF total tau and p-tau may provide additional information regarding neurodegeneration and neurofibrillary tangles load, respectively.38, 39 Therefore, our results support the concept that the interaction between brain amyloid-β plaques and neurofibrillary tangles drives downstream neurodegeneration in early disease stages. In addition, the better prediction obtained with CSF p-tau as compared with total tau might be explained by the possible higher specificity of p-tau to AD pathophysiology. Although high concentrations of p-tau better characterize the AD pathological process,40 abnormal CSF total tau may be found in several brain disorders associated with axonal death and neuronal loss.41 Indeed, two previous studies conducted in ADNI participants also found that CSF p-tau, but not total tau, modulates the link between amyloid-β pathology and brain atrophy or cognitive decline.12, 13 Although CSF amyloid-β1–42 is considered an effective biomarker for representing disease status, direct measurements of brain amyloid-β deposition, as measured with amyloid-PET imaging, might be more strongly associated with eminent disease progression in early AD stages.42, 43 Alternatively, the lack of association of CSF total tau and CSF amyloid-β1–42 with metabolic decline may be due to the characteristics of the participants of the study population or the CSF assay.
Clinical insights can be derived from the present results. For example, the combination of abnormalities of CSF p-tau and amyloid-PET biomarkers might serve as a strategy for population enrichment, whereas [18F]FDG PET may serve as a biomarker of efficacy in disease-modifying clinical trials that focus on individuals in the preclinical stages of AD.
The major methodological strength of the present results constitutes the use of continuous variables in our statistical analysis. In fact, biomarkers are naturally continuous measurements and dichotomization techniques, such as establishment of abnormalities thresholds have been constantly debated44 and might not be ideal way to evaluate a disease with a continuous spectrum as AD. Another strength in our analysis was the conservative multiple comparison correction threshold using a significance level of 0.001 to interpret the data, which helped to avoid false positives results. To the best of our knowledge, this is the first study showing the synergism between amyloidosis and neurodegeneration without assuming a threshold for biomarker abnormalities or artificial dichotomization techniques.
Methodological aspects limit the interpretation and external validity of the present results. It is important to mention that ADNI participants are highly educated; therefore mechanisms of brain and cognitive reserve may have a role in the [18F]FDG PET outcomes reported here.45 The study population is composed of a group of individuals who accepted to participate in a study focusing on AD. Therefore, our population may be a self-selected group of individuals who worry about cognition. It would be highly desirable to replicate these results in a larger population-based cohort. The observations reported here are purely phenomenological and do not intend to prove a biological synergy between amyloidosis and tau pathology. Our statistical model might have captured a sequence of events, involving amyloid-β aggregation, and subsequent tau phosphorylation and metabolic decline. Certainly, molecular biology approaches involving cell cultures and in vivo studies combining long-term sequential imaging of amyloid-β, tau and brain metabolism in animal models, and in humans could better assess a causal relation between these pathological proteins and brain metabolic decline. We presented the PET data without correcting for partial volume effects. Although partial volume correction had an impact on our SUVR values, this effect did not translate into significant differences in our final results. Previous studies suggest that hypometabolism is more related to APOE ɛ4 status than to amyloid-β accumulation.46 Interestingly, we found no interactive effects between APOE ɛ4 status and tau biomarkers, leading to hypometabolism. However, despite recent evidence suggesting no effect of APOE ɛ4 status in the metabolism of AD-related brain regions in ‘preclinical’ individuals,30 one should be cautious regarding the influence of APOE ɛ4 in the present results due to our relatively small sample size to detect genetic influences in early disease stages.
In conclusion, the proposed synergistic interaction between amyloid-β and tau supports the idea of a more integrative model of AD, although it does not explicitly refute the baptist or tauist perspectives.
References
Jack CR Jr, Knopman DS, Jagust WJ, Petersen RC, Weiner MW, Aisen PS et al. Tracking pathophysiological processes in Alzheimer's disease: an updated hypothetical model of dynamic biomarkers. Lancet Neurol 2013; 12: 207–216.
Hardy J, Selkoe DJ . The amyloid hypothesis of Alzheimer's disease: progress and problems on the road to therapeutics. Science 2002; 297: 353–356.
Jack CR Jr., Wiste HJ, Knopman DS, Vemuri P, Mielke MM, Weigand SD et al. Rates of beta-amyloid accumulation are independent of hippocampal neurodegeneration. Neurology 2014; 82: 1605–1612.
Knopman DS, Jack CR Jr., Wiste HJ, Weigand SD, Vemuri P, Lowe VJ et al. Selective worsening of brain injury biomarker abnormalities in cognitively normal elderly persons with beta-amyloidosis. JAMA Neurol 2013; 70: 1030–1038.
Mormino EC, Betensky RA, Hedden T, Schultz AP, Amariglio RE, Rentz DM et al. Synergistic effect of beta-amyloid and neurodegeneration on cognitive decline in clinically normal individuals. JAMA Neurol 2014; 71: 1379–1385.
Vos SJ, Xiong C, Visser PJ, Jasielec MS, Hassenstab J, Grant EA et al. Preclinical Alzheimer's disease and its outcome: a longitudinal cohort study. Lancet Neurol 2013; 12: 957–965.
Knopman DS, Jack CR Jr., Wiste HJ, Weigand SD, Vemuri P, Lowe V et al. Short-term clinical outcomes for stages of NIA-AA preclinical Alzheimer disease. Neurology 2012; 78: 1576–1582.
Roe CM, Fagan AM, Grant EA, Hassenstab J, Moulder KL, Maue Dreyfus D et al. Amyloid imaging and CSF biomarkers in predicting cognitive impairment up to 7.5 years later. Neurology 2013; 80: 1784–1791.
Duyckaerts C . Tau pathology in children and young adults: can you still be unconditionally baptist? Acta Neuropathol 2011; 121: 145–147.
Trojanowski JQ . Tauists, Baptists, Syners, Apostates, and new data. Ann Neurol 2002; 52: 263–265.
Fortea J, Vilaplana E, Alcolea D, Carmona-Iragui M, Sanchez-Saudinos MB, Sala I et al. Cerebrospinal fluid beta-amyloid and phospho-tau biomarker interactions affecting brain structure in preclinical Alzheimer disease. Ann Neurol 2014; 76: 223–230.
Desikan RS, McEvoy LK, Thompson WK, Holland D, Roddey JC, Blennow K et al. Amyloid-beta associated volume loss occurs only in the presence of phospho-tau. Ann Neurol 2011; 70: 657–661.
Desikan RS, McEvoy LK, Thompson WK, Holland D, Brewer JB, Aisen PS et al. Amyloid-beta—associated clinical decline occurs only in the presence of elevated P-tau. Arch Neurol 2012; 69: 709–713.
Ittner LM, Gotz J . Amyloid-beta and tau—a toxic pas de deux in Alzheimer's disease. Nat Rev Neurosci 2011; 12: 65–72.
Chabrier MA, Cheng D, Castello NA, Green KN, LaFerla FM . Synergistic effects of amyloid-beta and wild-type human tau on dendritic spine loss in a floxed double transgenic model of Alzheimer's disease. Neurobiol Dis 2014; 64: 107–117.
Khan UA, Liu L, Provenzano FA, Berman DE, Profaci CP, Sloan R et al. Molecular drivers and cortical spread of lateral entorhinal cortex dysfunction in preclinical Alzheimer's disease. Nat Neurosci 2014; 17: 304–311.
Jagust W, Reed B, Mungas D, Ellis W, Decarli C . What does fluorodeoxyglucose PET imaging add to a clinical diagnosis of dementia? Neurology 2007; 69: 871–877.
Ossenkoppele R, Tolboom N, Foster-Dingley JC, Adriaanse SF, Boellaard R, Yaqub M et al. Longitudinal imaging of Alzheimer pathology using [11C]PIB, [18F]FDDNP and [18F]FDG PET. Eur J Nucl Med Mol Imaging 2012; 39: 990–1000.
Dowling NM, Johnson SC, Gleason CE, Jagust WJ, Alzheimer's Disease Neuroimaging I. The mediational effects of FDG hypometabolism on the association between cerebrospinal fluid biomarkers and neurocognitive function. Neuroimage 2015; 105: 357–368.
Eskildsen SF, Coupe P, Fonov V, Manjon JV, Leung KK, Guizard N et al. BEaST: brain extraction based on nonlocal segmentation technique. Neuroimage 2012; 59: 2362–2373.
Mesulam MM . Neuroplasticity failure in Alzheimer's disease: bridging the gap between plaques and tangles. Neuron 1999; 24: 521–529.
Small SA, Duff K . Linking Abeta and tau in late-onset Alzheimer's disease: a dual pathway hypothesis. Neuron 2008; 60: 534–542.
Ittner LM, Ke YD, Delerue F, Bi M, Gladbach A, van Eersel J et al. Dendritic function of tau mediates amyloid-beta toxicity in Alzheimer's disease mouse models. Cell 2010; 142: 387–397.
Quintanilla RA, von Bernhardi R, Godoy JA, Inestrosa NC, Johnson GV . Phosphorylated tau potentiates Abeta-induced mitochondrial damage in mature neurons. Neurobiol Dis 2014; 71: 260–269.
Manczak M, Reddy PH . Abnormal interaction of oligomeric amyloid-beta with phosphorylated tau: implications to synaptic dysfunction and neuronal damage. J Alzheimer's Dis 2013; 36: 285–295.
Clinton LK, Blurton-Jones M, Myczek K, Trojanowski JQ, LaFerla FM . Synergistic Interactions between Abeta, tau, and alpha-synuclein: acceleration of neuropathology and cognitive decline. J Neurosci 2010; 30: 7281–7289.
Wang J, Yan K, Wu ZQ, Zheng CY, Xu RX, Chen LH et al. TDP-43 interaction with the intracellular domain of amyloid precursor protein induces p53-associated apoptosis. Neurosci Lett. 2014; 569: 131–136.
Kuhl DE, Metter EJ, Riege WH, Hawkins RA . The effect of normal aging on patterns of local cerebral glucose utilization. Ann Neurol 1984; 15: S133–S137.
Moeller JR, Ishikawa T, Dhawan V, Spetsieris P, Mandel F, Alexander GE et al. The metabolic topography of normal aging. J Cereb Blood Flow Metab 1996; 16: 385–398.
Lowe VJ, Weigand SD, Senjem ML, Vemuri P, Jordan L, Kantarci K et al. Association of hypometabolism and amyloid levels in aging, normal subjects. Neurology 2014; 82: 1959–1967.
Mosconi L, Mistur R, Switalski R, Brys M, Glodzik L, Rich K et al. Declining brain glucose metabolism in normal individuals with a maternal history of Alzheimer disease. Neurology 2009; 72: 513–520.
Thal DR, Rub U, Orantes M, Braak H . Phases of A beta-deposition in the human brain and its relevance for the development of AD. Neurology 2002; 58: 1791–1800.
Delacourte A, David JP, Sergeant N, Buee L, Wattez A, Vermersch P et al. The biochemical pathway of neurofibrillary degeneration in aging and Alzheimer's disease. Neurology 1999; 52: 1158–1165.
Braak H, Braak E . Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol 1991; 82: 239–259.
Sperling R, Mormino E, Johnson K . The evolution of preclinical Alzheimer's disease: implications for prevention trials. Neuron 2014; 84: 608–622.
Lesne SE, Sherman MA, Grant M, Kuskowski M, Schneider JA, Bennett DA et al. Brain amyloid-beta oligomers in ageing and Alzheimer's disease. Brain 2013; 136 (Pt 5): 1383–1398.
Ikonomovic MD, Klunk WE, Abrahamson EE, Mathis CA, Price JC, Tsopelas ND et al. Post-mortem correlates of in vivo PiB-PET amyloid imaging in a typical case of Alzheimer's disease. Brain 2008; 131 (Pt 6): 1630–1645.
Skillback T, Farahmand BY, Rosen C, Mattsson N, Nagga K, Kilander L et al. Cerebrospinal fluid tau and amyloid-beta1-42 in patients with dementia. Brain 2015; 138 (Pt 9): 2716–2731.
Blennow K, Hampel H, Weiner M, Zetterberg H . Cerebrospinal fluid and plasma biomarkers in Alzheimer disease. Nat Rev Neurol 2010; 6: 131–144.
Hampel H, Blennow K, Shaw LM, Hoessler YC, Zetterberg H, Trojanowski JQ . Total and phosphorylated tau protein as biological markers of Alzheimer's disease. Exp Gerontol 2010; 45: 30–40.
Brandt R, Hundelt M, Shahani N . Tau alteration and neuronal degeneration in tauopathies: mechanisms and models. Biochimi Biophys Acta 2005; 1739: 331–354.
Mattsson N, Insel PS, Donohue M, Landau S, Jagust WJ, Shaw LM et al. Independent information from cerebrospinal fluid amyloid-beta and florbetapir imaging in Alzheimer's disease. Brain 2015; 138 (Pt 3): 772–783.
Leuzy A, Carter SF, Chiotis K, Almkvist O, Wall A, Nordberg A . Concordance and diagnostic accuracy of [11C]PIB PET and cerebrospinal fluid biomarkers in a sample of patients with mild cognitive impairment and Alzheimer's Disease. J Alzheimers Dis 2015; 45: 1077–1088.
Villeneuve S, Rabinovici GD, Cohn-Sheehy BI, Madison C, Ayakta N, Ghosh PM et al. Existing Pittsburgh Compound-B positron emission tomography thresholds are too high: statistical and pathological evaluation. Brain 2015; 138: 2020–2033.
Stern Y . Cognitive reserve in ageing and Alzheimer's disease. Lancet Neurol 2012; 11: 1006–1012.
Jagust WJ, Landau SM, Alzheimer's Disease Neuroimaging I. Apolipoprotein E, not fibrillar beta-amyloid, reduces cerebral glucose metabolism in normal aging. J Neurosci 2012; 32: 18227–18233.
Acknowledgements
This work was supported by the Canadian Institutes of Health Research (CIHR; MOP-11-51-31), the Alan Tiffin Foundation, the Alzheimer's Association (NIRG-12-92090, NIRP-12-259245), Prevent-AD scholarship (TAP), Fonds de Recherche du Québec – Santé (FRQS; Chercheur Boursier, PR-N), S.G. and P.R-N. are members of the CIHR-CCNA Canadian Consortium of Neurodegeneration in Aging. Data collection and sharing for this project was funded by the Alzheimer’s Disease Neuroimaging Initiative (ADNI; National Institutes of Health Grant U01 AG024904) and DOD ADNI (Department of Defense award number W81XWH-12-2-0012). ADNI is funded by the National Institute on Aging, the National Institute of Biomedical Imaging and Bioengineering and through generous contributions from the following: AbbVie, Alzheimer’s Association; Alzheimer’s Drug Discovery Foundation; Araclon Biotech; BioClinica; Biogen; Bristol–Myers Squibb Company; CereSpir; Eisai Inc.; Elan Pharmaceuticals; Eli Lilly and Company; EuroImmun; F. Hoffmann-La Roche and its affiliated company Genentech; Fujirebio; GE Healthcare; IXICO; Janssen Alzheimer Immunotherapy Research & Development; Johnson & Johnson Pharmaceutical Research & Development; Lumosity; Lundbeck; Merck; Meso Scale Diagnostics; NeuroRx Research; Neurotrack Technologies; Novartis Pharmaceuticals Corporation; Pfizer; Piramal Imaging; Servier; Takeda Pharmaceutical Company; and Transition Therapeutics. The Canadian Institutes of Health Research is providing funds to support ADNI clinical sites in Canada. Private sector contributions are facilitated by the Foundation for the National Institutes of Health (www.fnih.org). The grantee organization is the Northern California Institute for Research and Education, and the study is coordinated by the Alzheimer's Disease Cooperative Study at the University of California, San Diego. ADNI data are disseminated by the Laboratory for Neuro Imaging at the University of Southern California.
Author information
Authors and Affiliations
Consortia
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
ALZHEIMER’S DISEASE NEUROIMAGING INITIATIVE
Data used in preparation of this article were obtained from the Alzheimer’s Disease Neuroimaging Initiative (ADNI) database (adni.loni.usc.edu). As such, the investigators within the ADNI contributed to the design and implementation of ADNI, and/or provided data, but did not participate in analysis or writing of this report. A complete listing of ADNI investigators can be found at: http://adni.loni.usc.edu/wp-content/uploads/how_to_apply/ADNI_Acknowledgement_List.pdf.
Supplementary Information accompanies the paper on the Molecular Psychiatry website
Supplementary information
PowerPoint slides
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Pascoal, T., Mathotaarachchi, S., Mohades, S. et al. Amyloid-β and hyperphosphorylated tau synergy drives metabolic decline in preclinical Alzheimer’s disease. Mol Psychiatry 22, 306–311 (2017). https://doi.org/10.1038/mp.2016.37
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mp.2016.37
This article is cited by
-
Cortical microstructural changes predict tau accumulation and episodic memory decline in older adults harboring amyloid
Communications Medicine (2023)
-
Neuroimaging of tissue microstructure as a marker of neurodegeneration in the AT(N) framework: defining abnormal neurodegeneration and improving prediction of clinical status
Alzheimer's Research & Therapy (2023)
-
Immunogenicity of MultiTEP platform technology-based Tau vaccine in non-human primates
npj Vaccines (2022)
-
[11C]Martinostat PET analysis reveals reduced HDAC I availability in Alzheimer’s disease
Nature Communications (2022)
-
Associations of AT(N) biomarkers with neuropsychiatric symptoms in preclinical Alzheimer’s disease and cognitively unimpaired individuals
Translational Neurodegeneration (2021)
