
Abstract
Enzymatic activity responsible for the cleavage of heparan sulfate, commonly known as heparanase, is abundant in tumor-derived cells. Heparanase cleaves heparan sulfate side chains, presumably at sites of low sulfation, thus facilitating structural alterations of the extracellular matrix and basement membrane underlying epithelial and endothelial cells. Traditionally, heparanase activity was correlated with the metastatic potential of tumor-derived cells, attributed to enhanced cell dissemination as a consequence of heparan sulfate cleavage and remodeling of the extracellular matrix barrier. More recently, heparanase upregulation was documented in an increasing number of human carcinomas and hematological malignancies, correlating with increased tumor metastasis, vascular density, and shorter post-operative survival of cancer patients. Although heparanase upregulation and its pro-malignant features are well documented, the instance of its induction in the course of tumor development was less investigated. Here, we used immunohistochemical analysis to investigate heparanase expression in normal esophagus, Barrett's esophagus without dysplasia, Barrett's esophagus with low-grade dysplasia, Barrett's esophagus with high-grade dysplasia, and adenocarcinoma of the esophagus. We report that heparanase expression is already induced in Barrett's epithelium without dysplasia, and is further increased during progression through distinct pathological stages, namely, low-grade dysplasia, high-grade dysplasia, and adenocarcinoma. Notably, heparanase induction correlated with increased cell proliferation index revealed by Ki-67 staining. These findings suggest that heparanase function is not limited to the process of tumor metastasis, but rather is engaged at the early stages of esophagus carcinoma initiation and progression.
Similar content being viewed by others

Main
Heparanase is an endo-β-D-glucuronidase, the predominant enzyme that degrades heparan sulfate side chains of heparan sulfate proteoglycans.1, 2 These complex macromolecules are highly abundant in the extracellular matrix and are thought to have an important structural role, contributing to extracellular matrix integrity and insolubility.3, 4 Traditionally, heparanase activity was correlated with the metastatic potential of tumor-derived cells, attributed to enhanced cell dissemination as a consequence of heparan sulfate cleavage and remodeling of the extracellular matrix barrier.1, 2 A proof-of-concept to this notion has been established by using specific anti-heparanase ribozyme and siRNA methodologies, clearly implicating heparanase-mediated heparan sulfate cleavage as a critical requisite for metastatic spread.5 Similarly, heparanase activity was implicated in cell dissemination associated with inflammation and angiogenesis.5, 6 More recently, heparanase upregulation was documented in an increasing number of human carcinomas and hematological malignancies.7, 8 In many cases, heparanase induction correlated with increased tumor metastasis, vascular density, and shorter post-operative survival of cancer patients, thus providing a strong clinical support for the pro-metastatic and pro-angiogenic functions of the enzyme and encouraging the development of heparanase inhibitors such as anti-cancer drugs.9, 10, 11, 12
Heparanase expression by the gastrointestinal tract has been studied employing immunostaining, RT-PCR, and enzymatic activity analyses. In these studies, heparanase was not detected in normal-looking epithelia, whereas high levels of expression were observed in colon and gastric carcinomas.13, 14, 15, 16, 17
Heparanase induction has similarly been observed in carcinoma of the esophagus, and its expression correlated with tumor malignancy and poor survival.18, 19, 20, 21 Although heparanase upregulation and its pro-malignant features are well documented, the timing of its induction in the course of tumor development is less investigated and understood. Barrett's esophagus is defined as the replacement (metaplasia) of the normal esophageal squamous mucosa with columnar epithelium.22, 23 This condition most commonly arises in the setting of chronic gastroesophageal reflux disease, in which repeated mucosal injury is believed to stimulate metaplasia. The abnormal Barrett's epithelium predisposes patients to the development of epithelial dysplasia and, ultimately, to adenocarcinoma.22, 23 Adenocarcinomas seem to progress in a step-wise manner, from low-grade to high-grade dysplasia, and to intramucosal and finally to invasive carcinoma. Here, we used immunohistochemistry analysis to investigate heparanase expression in normal esophagus, Barrett's esophagus without dysplasia, Barrett's esophagus with low-grade dysplasia, Barrett's esophagus with high-grade dysplasia, and adenocarcinoma of the esophagus.
Materials and methods
Experimental Design
The study included archived paraffin-embedded esophageal sections obtained from 66 patients who underwent esophagogastroduodenoscopy with biopsy or surgical esophageal resection. The cohort included 11 samples of normal esophagus, 21 Barrett's esophagus without dysplasia, 5 Barrett's esophagus with low-grade dysplasia, 12 Barrett's esophagus with high-grade dysplasia, and 17 adenocarcinomas, diagnosed at the Rambam Health Care Campus, Haifa, and the Hillel-Yaffe Medical Center, Hadera, Israel. The grade of dysplasia was defined according to the degree of nuclear atypia and glandular architectural changes.24 Paraffin sections were stained with anti-heparanase antibodies and heparanase staining intensity was correlated to pathological staging and proliferation index revealed by anti-Ki-67 staining. The study protocol was approved by the Institutional Review Board.
Immunostaining
Staining of formalin-fixed, paraffin-embedded 5-μm sections for heparanase was performed essentially as described.25, 26 Briefly, slides were deparaffinized, rehydrated, and endogenous peroxidase activity was quenched (30 min) by 0.3% hydrogen peroxide in methanol. Slides were then subjected to antigen retrieval by boiling (20 min) in 10 mM citrate buffer, pH 6. Slides were incubated with 10% normal goat serum in phosphate-buffered saline (PBS) for 60 min to block non-specific binding, followed by incubation (20 h, 4°C) with polyclonal anti-heparanase 733 antibodies,27 diluted 1:100 in blocking solution. Slides were extensively washed with PBS containing 0.01% Triton X-100 and incubated with a secondary reagent (Envision kit) according to the manufacturer's (Dako, Glostrup, Denmark) instructions. After additional washes, color was developed using the AEC reagent (Dako), sections were counterstained with hematoxylin and mounted, as described.25, 26 Immunostained specimens were examined by a senior pathologist and were scored according to the intensity of staining (0=none; 1=weak to moderate; and 2=strong). In addition, staining localization was classified as nuclear (n), cytoplasmic (c), or both (n+c). Slides were similarly stained with anti-Ki-67 monoclonal antibody (MIB-1; Dako). Biotinylated secondary antibody was applied for 10 min, followed by incubation with streptavidin peroxidase (Dako) for 10 min. Slides were then washed extensively and color was developed using AEC reagent (Dako) and counterstained with hematoxylin as above. Cell proliferation index was assessed by a senior pathologist and categorized as low (<5%), moderate (6–50%), and high (>50%) according to the percentage of Ki-67-positive cells.
Statistical Analysis
The data were evaluated by SPSS software, version 15 (SPSS, Chicago, IL, USA). Fisher's Exact Test and Pearson's χ2-test were used for detection of differences in the prevalence of the staining intensity between different diagnoses. Differences between heparanase intensity and the percentage of Ki-67 staining were assessed by non-parametric Mann–Whitney U-test. P<0.05 was considered significant.
Results
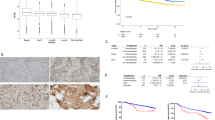
Heparanase staining was not detected in normal esophageal epithelium (Table 1, Figure 1a). In contrast, positive staining of heparanase was observed in 52% (11/21) of the patients diagnosed with Barrett's esophagus without dysplasia (Figure 1b; Table 1), an increase that is statistically highly significant (P=0.0045). In Barrett's esophagus, heparanase was mainly confined to intestinal-type epithelium and was localized at the basal aspect of the epithelium in a spotted manner (Figure 1b). Gastric–cardiac epithelium and gastric body-type epithelium often mingled with intestinal epithelium of Barrett's esophagus, and rarely stained for heparanase. Further enhancement in heparanase staining was observed in high-grade dysplasia (P=0.005; Figure 1d), wherein all cases stained positively (Table 1). Similarly, all cases diagnosed as esophagus adenocarcinoma stained positive for heparanase (Table 1; Figure 1e). Moreover, although in high-grade dysplasia 33% of the cases (4/12) exhibited strong (+2) staining of heparanase, 88% (15/17) of the adenocarcinoma biopsies exhibited such staining (P=0.0045) (Table 1). Unlike the spotted appearance noted in Barrett's esophagus (Figure 1b) and low-grade dysplasia (Figure 1c), heparanase staining appeared diffuse throughout the cell cytoplasm of high-grade dysplasia (Figure 1d) and adenocarcinoma (Figure 1e) specimens. Careful examination revealed further differences in heparanase cellular localization. Thus, although in high-grade specimens heparanase staining appeared mostly cytoplasmic (Figure 1d; Figure 2a), 71% (12/17) of the adenocarcinoma cases also exhibited nuclear localization of heparanase (Figure 2b) compared with only one case of high-grade dysplasia exhibiting such localization (P=0.0018) (Figure 2c).

Immunohistochemical staining of esophageal specimens. Heparanase staining. Formalin-fixed, paraffin-embedded 5-μm sections of normal (a), Barrett's (b), low-grade (c), high-grade (d), and adenocarcinoma (e) esophageal biopsies were subjected to immunostaining of heparanase, applying anti-heparanase polyclonal Antibody no 733, as described under ‘Materials and Methods’. Specimens that were similarly stained applying the above procedure, but lacking the primary antibody, yielded no detectable staining (f, adenocarcinoma). Original magnification: (a) × 200; (b–f) × 400.

Heparanase cellular localization. Shown are representative photomicrographs of cytoplasmic (a) and nuclear (b) staining of heparanase in esophageal adenocarcinoma. Original magnification: × 400. Distribution (percent) of cases exhibiting cytoplasmic vs nuclear localization of heparanase in high-grade dysplasia and adenocarcinoma is presented graphically in (c).
To further elucidate the clinical significance of heparanase expression in esophagus cancer, cell proliferation was evaluated by staining for Ki-67. As expected, a low rate of cell proliferation was noted in normal and Barrett's epithelium (Figure 3a, b, respectively). In Barrett's esophagus, Ki-67 staining primarily decorated the intestinal epithelium. Cell proliferation was markedly increased in high-grade dysplasia (Figure 3d) and adenocarcinoma (Figure 3e). Significantly, heparanase staining correlated with cell proliferation index (P=0.0006) (Table 2). Thus, although 87% of the cases that were stained negative for heparanase exhibited a low (<5) proliferative index, most heparanase-positive cases (70%) showed medium to high proliferation rate (Table 2). These findings imply that heparanase may have a role in esophageal cancer initiation and progression.

Immunohistochemical Ki-67 staining of esophageal specimens. Formalin-fixed, paraffin-embedded 5-μm sections of normal (a), Barrett's (b), low-grade (c), high-grade (d), and adenocarcinoma (e) esophageal biopsies were subjected to immunostaining of Ki-67 as described under ‘Materials and Methods’. Original magnification: × 200.
Discussion
It is now generally accepted that Barrett's epithelium can progress through a metaplasia–dysplasia–carcinoma sequence.22, 23 Barrett's epithelium is, therefore, considered a premalignant condition with a 50–100-fold increased risk of cancer.22, 23 The etiology of malignant Barrett's esophagus remains largely unknown and is likely multifactorial. Previous studies showed that heparanase expression is induced in carcinoma of the esophagus.18, 20 Heparanase induction correlated with enhanced tumor cell invasion into the muscular and adventitia layers, increased tumor metastasis, and advanced tumor stage, altogether associated with poor survival.18, 20 High heparanase staining in esophageal adenocarcinoma biopsies noted in this study (Table 1) further supports this notion. Interestingly, however, increased heparanase expression was already noted in Barrett's epithelium, representing the earliest stage of esophagus carcinogenesis (Table 1; Figure 1). Further increase in heparanase expression paralleled the progression through distinct pathological stages, namely, low-grade dysplasia, high-grade dysplasia, and adenocarcinoma (Table 1, Figure 1). These findings suggest that heparanase function is not limited to the process of tumor metastasis, but rather is engaged at the early stages of esophagus carcinoma initiation and progression to adenocarcinoma. Intimate involvement in esophageal cancer development is further supported by the close association found between heparanase expression and cell proliferation. Clearly, biopsies stained positively for heparanase exhibited a higher proliferative index than did heparanase-negative specimens (Table 2). Reduced proliferation rates noted after heparanase gene silencing in tumor-derived cells28 supports this notion and suggest that its pro-proliferative function is a common feature. Thus, heparanase inhibitors, such as non-anticoagulant glycol-split heparin12, 29 may be considered as a prophylactic treatment for Barrett's patients. This possibility is supported by recent studies showing no adverse side effects in mice treated with this drug for a relatively long period of time,30 yet it awaits in-depth preclinical and clinical evaluation. Whether heparanase plasma levels could serve as a biomarker for distinguishing Barrett's cases that are more likely to progress into adenocarcinoma is yet to be revealed and requires long-term follow-up and a large cohort of patients. A recently developed heparanase ELISA assay 31, 32, 33 may be applied to monitor heparanase levels in Barrett's patients. This approach would not only provide another indication for heparanase upregulation and secretion into the circulation but may also ideally substitute for the invasive endoscopic surveillance currently employed to monitor Barret's patients.
Esophagus cancer progression was also associated with alterations in heparanase cellular localization. In Barrett's and low-grade dysplasia, heparanase appeared punctuated and was localized to the basal aspect of the epithelium (Figure 1b, c). Interestingly, a similar staining pattern was noted in adenoma of the colon and in inflammatory bowel disease specimens,14, 26 both representing premalignant conditions. In contrast, heparanase appeared more diffusely distributed in the cell cytoplasm of high-grade dysplasia (Figure 1d) and adenocarcinoma specimens (Figure 1e) representing, possibly, heparanase protein that is more readily secreted. This is in agreement with the elevation of heparanase levels in the plasma and urine of cancer patients.31, 32, 33 The significance of this observation critically emerges from recent studies showing that secreted heparanase augments signal transduction pathways, leading to activation of certain protein kinases (that is, Akt, Src, EGFR) and increased gene transcription (that is, VEGF, TF, COX2) associated with enhanced tumor progression.21, 28, 34, 35, 36, 37, 38, 39
Previous studies have shown that heparanase also assumes nuclear localization,40, 41 and that nuclear heparanase is associated with a favorable prognosis of head and neck, gastric, and esophageal cancer patients.20, 41, 42, 43 It was unclear, however, at what stage of tumor initiation/progression nuclear translocation takes place. Likewise, nuclear localization of heparanase was observed in the present study (Figure 2b, c), supporting its prevalence in carcinoma of the esophagus. Notably, nuclear localization was observed primarily in esophagus adenocarcinoma, and only occasionally in high-grade dysplasia (Figure 2c). These findings suggest that nuclear translocation of heparanase takes place mainly at the final stages of tumor progression into adenocarcinoma. Identifying the molecular mechanism underlying this process and the associated favorable outcome is a major challenge for future research focusing on heparanase.
References
Parish CR, Freeman C, Hulett MD . Heparanase: a key enzyme involved in cell invasion. Biochim Biophys Acta 2001;1471:M99–108.
Vlodavsky I, Friedmann Y . Molecular properties and involvement of heparanase in cancer metastasis and angiogenesis. J Clin Invest 2001;108:341–347.
Bishop JR, Schuksz M, Esko JD . Heparan sulphate proteoglycans fine-tune mammalian physiology. Nature 2007;446:1030–1037.
Iozzo RV . Basement membrane proteoglycans: from cellar to ceiling. Nat Rev Mol Cell Biol 2005;6:646–656.
Edovitsky E, Elkin M, Zcharia E, et al. Heparanase gene silencing, tumor invasiveness, angiogenesis, and metastasis. J Natl Cancer Inst 2004;96:1219–1230.
Edovitsky E, Lerner I, Zcharia E, et al. Role of endothelial heparanase in delayed-type hypersensitivity. Blood 2006;107:3609–3616.
Ilan N, Elkin M, Vlodavsky I . Regulation, function and clinical significance of heparanase in cancer metastasis and angiogenesis. Int J Biochem Cell Biol 2006;38:2018–2039.
Vreys V, David G . Mammalian heparanase: what is the message? J Cell Mol Med 2007;11:427–452.
Ferro V, Hammond E, Fairweather JK . The development of inhibitors of heparanase, a key enzyme involved in tumour metastasis, angiogenesis and inflammation. Mini Rev Med Chem 2004;4:693–702.
McKenzie EA . Heparanase: a target for drug discovery in cancer and inflammation. Br J Pharmacol 2007;151:1–14.
Miao HQ, Liu H, Navarro E, et al. Development of heparanase inhibitors for anti-cancer therapy. Curr Med Chem 2006;13:2101–2111.
Vlodavsky I, Ilan N, Naggi A, et al. Heparanase: structure, biological functions, and inhibition by heparin-derived mimetics of heparan sulfate. Curr Pharm Des 2007;13:2057–2073.
Chen JQ, Zhan WH, He YL, et al. Expression of heparanase gene, CD44v6, MMP-7 and nm23 protein and their relationship with the invasion and metastasis of gastric carcinomas. World J Gastroenterol 2004;10:776–782.
Doviner V, Maly B, Kaplan V, et al. Spatial and temporal heparanase expression in colon mucosa throughout the adenoma-carcinoma sequence. Mod Pathol 2006;19:878–888.
Friedmann Y, Vlodavsky I, Aingorn H, et al. Expression of heparanase in normal, dysplastic, and neoplastic human colonic mucosa and stroma. Evidence for its role in colonic tumorigenesis. Am J Pathol 2000;157:1167–1175.
Nobuhisa T, Naomoto Y, Ohkawa T, et al. Heparanase expression correlates with malignant potential in human colon cancer. J Cancer Res Clin Oncol 2005;131:229–237.
Tang W, Nakamura Y, Tsujimoto M, et al. Heparanase: a key enzyme in invasion and metastasis of gastric carcinoma. Mod Pathol 2002;15:593–598.
Mikami S, Ohashi K, Katsube K, et al. Coexpression of heparanase, basic fibroblast growth factor and vascular endothelial growth factor in human esophageal carcinomas. Pathol Int 2004;54:556–563.
Mikami S, Ohashi K, Usui Y, et al. Loss of syndecan-1 and increased expression of heparanase in invasive esophageal carcinomas. Jpn J Cancer Res 2001;92:1062–1073.
Ohkawa T, Naomoto Y, Takaoka M, et al. Localization of heparanase in esophageal cancer cells: respective roles in prognosis and differentiation. Lab Invest 2004;84:1289–1304.
Okawa T, Naomoto Y, Nobuhisa T, et al. Heparanase is involved in angiogenesis in esophageal cancer through induction of cyclooxygenase-2. Clin Cancer Res 2005;11:7995–8005.
Quinlan JM, Colleypriest BJ, Farrant M, et al. Epithelial metaplasia and the development of cancer. Biochim Biophys Acta 2007;1776:10–21.
Schuchert MJ, Luketich JD . Barrett's esophagus-emerging concepts and controversies. J Surg Oncol 2007;95:185–189.
Odze RD . Diagnosis and grading of dysplasia in Barrett's oesophagus. J Clin Pathol 2006;59:1029–1038.
Cohen E, Doweck I, Naroditsky I, et al. Heparanase is overexpressed in lung cancer and correlates inversely with patient survival. Cancer 2008;113:1004–1011.
Waterman M, Ben-Izhak O, Eliakim R, et al. Heparanase upregulation by colonic epithelium in inflammatory bowel disease. Mod Pathol 2007;20:8–14.
Zetser A, Levy-Adam F, Kaplan V, et al. Processing and activation of latent heparanase occurs in lysosomes. J Cell Sci 2004;117:2249–2258.
Cohen-Kaplan V, Doweck I, Naroditsky I, et al. Heparanase augments epidermal growth factor receptor phosphorylation: correlation with head and neck tumor progression. Cancer Res 2008;68:10077–10085.
Casu B, Vlodavsky I, Sanderson RD . Non-anticoagulant heparins and inhibition of cancer. Pathophysiol Haemost Thromb 2008;36:195–203.
Yang Y, MacLeod V, Dai Y, et al. The syndecan-1 heparan sulfate proteoglycan is a viable target for myeloma therapy. Blood 2007;110:2041–2048.
Shafat I, Ben-Barak A, Postovsky S, et al. Heparanase levels are elevated in the plasma of pediatric cancer patients and correlate with response to anticancer treatment. Neoplasia 2007;9:909–916.
Shafat I, Pode D, Peretz T, et al. Clinical significance of urine heparanase in bladder cancer progression. Neoplasia 2008;10:125–130.
Shafat I, Zcharia E, Nisman B, et al. An ELISA method for the detection and quantification of human heparanase. Biochem Biophys Res Commun 2006;341:958–963.
Ben-Zaken O, Gingis-Velitski S, Vlodavsky I, et al. Heparanase induces Akt phosphorylation via a lipid raft receptor. Biochem Biophys Res Commun 2007;361:829–834.
Cohen-Kaplan V, Naroditsky I, Zetser A, et al. Heparanase induces VEGF C and facilitates tumor lymphangiogenesis. Int J Cancer 2008;123:2566–2573.
Fux L, Feibish N, Cohen-Kaplan V, et al. Structure-function approach identifies a C-terminal domain that mediates heparanase signaling. Cancer Res 2009;69:1758–1767.
Gingis-Velitski S, Zetser A, Flugelman MY, et al. Heparanase induces endothelial cell migration via protein kinase B/Akt activation. J Biol Chem 2004;2004:279:23536–279:23541.
Nadir Y, Brenner B, Zetser A, et al. Heparanase induces tissue factor expression in vascular endothelial and cancer cells. J Thromb Haemost 2006;4:2443–2451.
Zetser A, Bashenko Y, Edovitsky E, et al. Heparanase induces vascular endothelial growth factor expression: correlation with p38 phosphorylation levels and Src activation. Cancer Res 2006;66:1455–1463.
Schubert SY, Ilan N, Shushy M, et al. Human heparanase nuclear localization and enzymatic activity. Lab Invest 2004;84:535–544.
Takaoka M, Naomoto Y, Ohkawa T, et al. Heparanase expression correlates with invasion and poor prognosis in gastric cancers. Lab Invest 2003;83:613–622.
Doweck I, Kaplan-Cohen V, Naroditsky I, et al. Heparanase localization and expression by head and neck cancer: correlation with tumor progression and patient survival. Neoplasia 2006;8:1055–1061.
Kobayashi M, Naomoto Y, Nobuhisa T, et al. Heparanase regulates esophageal keratinocyte differentiation through nuclear translocation and heparan sulfate cleavage. Differentiation 2006;74:235–243.
Acknowledgements
This work was supported by grants from the Israel Science Foundation (grant 549/06); National Cancer Institute, NIH (grant RO1-CA106456); and the DKFZ-MOST cooperation program in cancer research. Israel Vlodavsky is a Research Professor of the Israel Cancer Research Fund (ICRF).
Author information
Authors and Affiliations
Corresponding author
Additional information
Disclosure/conflict of interest
The authors declare no conflict of interest.
Rights and permissions
About this article
Cite this article
Brun, R., Naroditsky, I., Waterman, M. et al. Heparanase expression by Barrett's epithelium and during esophageal carcinoma progression. Mod Pathol 22, 1548–1554 (2009). https://doi.org/10.1038/modpathol.2009.115
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/modpathol.2009.115
Keywords
This article is cited by
-
Parabacteroides produces acetate to alleviate heparanase-exacerbated acute pancreatitis through reducing neutrophil infiltration
Microbiome (2021)
-
Heparanase and the hallmarks of cancer
Journal of Translational Medicine (2020)
-
Significance of Heparanase in Cancer and Inflammation
Cancer Microenvironment (2012)
