
Abstract
Anti-adhesion therapies that target α4 integrins (e.g., natalizumab) are thought to work by blocking T-cell recruitment to the intestinal tissues in patients with Crohn’s disease (CD); however, little direct evidence is available to confirm this contention. We wished to evaluate the importance of T cell-associated α4 integrins in a chronic colitis model in mice and to determine the effect of natalizumab treatment on intestinal tissue T-cell accumulation in human CD. Adoptive transfer of T cells lacking α4 (α4−/−) but not β1 integrin into immunodeficient mice produced significantly attenuated disease. This was correlated with reduced numbers of colon CD4 T cells compared with the control mice; however, tissue distribution of T helper type 1 (Th1) and T helper type 17 (Th17) cells and regulatory T cells (Tregs) was not affected by the lack of α4. Furthermore, α4−/− T cells demonstrated defective homing to the chronically inflamed small intestines and colons. Finally, patients treated with natalizumab showed significant reduction in mucosal CD4 T cells and no skewing in the foxp3+ Treg or T-bet+Th1 fractions thereof. These results demonstrate a direct role for T cell-associated α4β7 but not α4β1 integrins during initiation and perpetuation of chronic colitis. Moreover, our data demonstrated that natalizumab treatment reduced mucosal CD4 T-cell accumulation in CD patients.
Similar content being viewed by others

INTRODUCTION
Development of Crohn’s disease (CD) is associated with the accumulation of immune cells in the gastrointestinal tract. Emerging experimental and clinical evidence suggests that leukocyte-associated α4 integrins may play an important role in the recruitment of these cells to the intestinal tissues, thus contributing to induction and perpetuation of chronic intestinal inflammation.1, 2, 3, 4, 5, 6 The α4 integrins belong to a family of heterodimeric proteins that mediate adhesive and signaling interactions between circulating leukocytes and endothelial cells. The α4 chain can combine with either β1 or β7 chains to form very late antigen-4 (α4β1) or lymphocyte Peyer’s patch adhesion molecule 1 (LPAM-1; α4β7) heterodimers, respectively.
Investigations into the roles of T cell-associated α4 integrins in chronic mouse models of intestinal inflammation have been difficult to perform because of the embryonic lethality of the α4-deficiency in mice.7 To circumvent this, β7-deficient (β7−/−) mice were utilized to address the relevance of α4β7 and αEβ7 in experimental colitis. However, it produced contrasting results, with some studies demonstrating that adoptive transfer of β7−/− T cells into the immunodeficient recipients delayed the onset but not the severity of colitis,8, 9 whereas others showed attenuated disease.3 Other indirect methods have been used to assess the role of α4 integrins in the pathogenesis of experimental inflammatory bowel disease (IBD). For example, Picarella et al.10 found that treatment of colitic mice with antibodies to β7 integrin and mucosal addressin cell adhesion molecule-1 (MadCAM-1) attenuated disease development. Rivera-Nieves et al.6 demonstrated that antibody-mediated blockade of either α4β1 or α4β7 individually did not attenuate chronic ileitis in SAMP1/Yit mice, whereas immune neutralization of both (or their endothelial ligands) provided significant protection. On the other hand, at least one study found that long-term administration of anti-α4 integrin monoclonal antibody exacerbated colonic inflammation in Gαi2-deficient mice.11 Moreover, studies in cotton-top tamarin monkeys that spontaneously develop IBD-like disease in captivity revealed that administration of anti-α4 (see ref 4) or anti-α4β7 monoclonal antibodies12 led to significant attenuation of their disease, suggesting a critical role of α4 in the development of IBD.
Targeting leukocyte recruitment from blood into intestine has been successfully developed into a therapy for moderate to severe CD. Natalizumab is a monoclonal antibody that targets the α4 integrin, thus blocking α4β1 and α4β7 molecules. Several clinical trials demonstrated the effectiveness of natalizumab at increasing the rates of clinical remission and the response rates2, 13 or its efficacy as induction and maintenance therapy in a subset of CD patients who initially responded to this treatment.14 Another anti-adhesion therapy is vedolizumab, a humanized antibody to α4β7. Recently completed phase 3 clinical trials demonstrated that vedolizumab was effective at treating ulcerative colitis and CD patients.15, 16
The protective results obtained using monoclonal antibodies directed against the α4 integrin or α4β7 has been used as evidence to implicate the importance of T cells in the induction or perpetuation of disease. However, T cells are a diverse and multifunctional population that may not be uniformly affected by α4 blockade. In the blood, expression of integrins α4 and β7 is markedly lower on Foxp3+ regulatory T cells (Tregs) than conventional CD4+ T cells,17 despite Tregs being present in abundance in the intestinal mucosa.18 In addition, in vivo administration of monoclonal antibodies may affect other cells. For example, α4β7 is found on B cells, natural killer cells, monocytes, and eosinophils, whereas α4β1 is expressed by neutrophils.19, 20, 21 Although α4β1 is not important in mediating T-cell recruitment to the gastrointestinal tract under steady-state noninflammatory conditions, in active CD, upregulation of its ligand, vascular cell adhesion molecule-1 (VCAM-1), by endothelial cells of the intestine22 may contribute to leukocyte recruitment into the inflamed gut via α4β1/VCAM-1.23, 24
Therefore, the objectives of this study were to ascertain the relative importance of T cell-associated α4 integrins, namely α4β7 and α4β1, in the induction of chronic gut inflammation in mice and to evaluate the importance of α4 integrins for mucosal T-cell accumulation in human CD. To accomplish the first objective, we generated conditional mutant mice that selectively lack the T cell-associated gene α4itg or β1itg and evaluated the ability of α4−/− and β1−/− T cells to induce chronic colitis using T-cell transfer model of colitis. We found that deletion of α4 integrin but not β1 integrin in T cells significantly attenuated development of colitis in mice and reduced accumulation of T cells in the colons. These findings in mice also correlated with the reduced accumulation of CD4 T cells in the intestinal biopsies of human CD patients treated with natalizumab. Taken together, our results demonstrated a critical role for T cell-associated α4 integrins in the induction and the perpetuation of CD and suggested a multifaceted role of this molecule in T-cell biology.
RESULTS
Adoptive transfer of CD45RBhigh T cells lacking surface expression of α4 integrin produced attenuated colitis in the immunodeficient recipient mice
To define the role of α4 integrins in the pathogenesis of chronic colitis, we generated a conditional knockout mouse using cre-loxP-mediated recombination technology using two complementary approaches. For the first approach, we injected polyinosinic:polycytidylic acid (poly I:C) into Mx.cre+α4loxP/loxP (Mx.cre+) mice that resulted in the loss of α4 integrin in 40–70% of CD4+ T cells (Supplementary Figure S1A online). Lack of α4 integrin expression did not affect their in vitro proliferation (Supplementary Figure S1B). To evaluate colitogenic potential of these cells, CD4+CD45RBhigh T cells from polyI:C-injected Mx.cre+ mice or Mx.cre-negative mice were purified by fluorescence-activated cell sorting into α4+ and α4neg subsets and adoptively transferred into recombination activating gene-1-deficient (RAG-1−/−) mice. Whereas transfer of α4+ T cells induced severe colitis, α4neg T cells produced little or no disease (Supplementary Figure S1E). Significantly fewer CD4 T cells were isolated from the colons of α4neg→RAG-1−/− mice compared with α4+→ RAG-1−/− mice (Supplementary Figure S1F). To our surprise, analysis of surface integrin molecules on CD4+ T cells in the reconstituted α4neg→RAG-1−/− mice revealed re-expression of α4 integrin even despite our most stringent gating during the sort (Supplementary Figure S2). These data suggested that α4 integrins are important for the T cell-induced chronic colitis, although cell sorting approach was insufficient to obtain a pure population of α4neg T cells.
Mice with T cell-specific deletion of α4 show normal levels of T cells in their intestine and gut-associated lymphoid tissue (GALT)
In order to delete itgα4 gene in T cells, we crossed α4loxP/loxP mice with CD4-cre mice. In the α4loxP/loxPCD4Cre+ (hereafter α4−/−) mice, α4 was not detected on the surface of CD4 and CD8 T cells and in ∼40% of B cells (Figure 1a). At the same time, expression of other major adhesion molecules CD29 (β1 integrin), CD11a (lymphocyte function-associated antigen-1 (LFA-1), αL integrin), CD18 (β2 integrin), and CD54 (intercellular adhesion molecule 1 (ICAM-1)) was comparable to littermate control α4loxP/loxPCD4Cre- (α4+/+) mice (Supplementary Figure S3).

CD4+CD45RBhigh T cells lacking surface expression of α4 integrins induce significantly less colitis than their wild-type (WT) counterparts following their transfer into recombination activating gene-1-deficient (RAG-1−/−) mice. (a) Specific deletion of α4 in T cells in α4−/− (α4f/fxCD4Cre+) but not α4+/+ (α4f/fxCD4Cre−). Representative histograms are shown. Isotype control antibody staining is indicated by tinted gray plots. (b) Blinded histopathological distal colon scores and (c) representative hematoxylin and eosin (H&E)-stained images that were taken at × 100 magnification. (d) Absolute numbers of CD4+ T cells. Data are presented as mean values±s.e.m. for 12–14 mice analyzed per group across at least three independent experiments. (e) Plasma levels of inflammatory cytokines. Scatter plots with mean values per group (black lines) are shown. Samples from α4+/+→RAG-1−/− and WT→RAG-1−/− mice were combined as “α4+/+”. Significant differences are indicated as*P<0.05 and ***P<0.001.
Deficiency in β7 integrin leads to a severe impairment in GALT formation and reduced numbers of intestinal lymphocytes.25 Thus, we were somewhat surprised to find that α4−/− mice showed comparable number and structure of Peyer’s patches compared with control mice (Supplementary Figure S4A online). Moreover, CD4 and CD8 T-cell distribution in the intestine and the GALT were similar between the two groups (Supplementary Figure S4B,C online). To determine whether lack of α4 may affect some intestinal T-cell subsets more than others without changing the overall number, we carried out differential analysis of intestinal and GALT T-cell receptor-αβ and -γδ T cells. We did not find any significant alterations in T-cell numbers between α4−/− and α4+/+ mice (Supplementary Figure S5).
The α4-deficient CD45RBhigh T cells are significantly attenuated in their ability to induce chronic colitis
Adoptive transfer of α4−/− CD4+CD45RBhigh T cells into RAG-1−/− mice produced significantly attenuated colitis compared with those that received α4+/+ T cells or T cells isolated from α4wt/wt CD4.Cre+ mice (Figure 1b,c). Flow cytometry analysis revealed comparable accumulation of CD4 T cells in the spleens of α4−/−→ RAG-1−/− and α4+/+→ RAG-1−/− mice, whereas their numbers in mesenteric lymph nodes (MLNs) and colonic lamina propria (LP) were significantly reduced (Figure 1d). Blood level of interferon-γ (IFN-γ) was significantly reduced in the noncolitic α4−/−→ RAG-1−/− mice, whereas levels of interleukin (IL)-12p70, IL-17, and tumor necrosis factor-α were comparable to those with severe disease (Figure 1e).
Expression of surface adhesion molecules on T cells isolated from the recipients’ intestinal tissues was comparable between the two groups (Figure 2a,b). Interestingly, expression of α4β7 integrin in the α4+/+→ RAG-1−/− group was absent from the surface of T cells isolated from small intestine and colon LP, but was seen on T cells from MLNs (Figure 2a), suggesting that it is downregulated following entry of T cells into the colon LP. Expression of α4 or β7 integrins paralleled tissue-distribution pattern that was seen with α4 and α4β7 staining. Expression of β7 integrin on some T cells in the α4−/−→ RAG-1−/− group was because of pairing with CD103 (αE integrin). Noteworthy, colon LP T cells in both groups showed prominent expression of β1 integrin (Figure 2a). Lastly, T cells in both groups displayed high levels of expression of CD44 and lacked expression of CD62L (data not shown) confirming that they had acquired activated/effector phenotype.

Expression of α4 integrins on the surface of wild-type (WT) CD4 T cells from colitic mice. (a) Representative histograms showing staining with specific antibodies (black lines) or isotype control antibodies (gray tinted lines) were initially gated on CD4+ T cells. Numbers show percent of positive T cells. (b) Combined data from three mice per group are expressed as mean±s.e.m.
Lack of α4 integrin does not affect in vitro polarization and tissue accumulation of Treg, Th1, or Th17 cells
Recent reports found that expression of α4 integrins is important for recruitment of T helper type 1 (Th1) cells into the brain and central nervous system during the development of experimental autoimmune encephalomyelitis.26, 27 Lack of α4 integrin did not affect polarization of T cells into Th1, Th2, or Th17 subsets (Supplementary Figure S6A).
In agreement with previous reports,26, 27 we confirmed higher expression of α4 integrin on the surface of Th1 cells as compared with Th17 cells (Supplementary Figure S6B). Therefore, we next wished to determine whether transfer of α4−/− T cells produced an imbalance between Th1 and Th17 cells in colitic mice. Despite significantly fewer T cells in the colon LP of the α4−/−→ RAG-1−/− group, intracellular staining revealed a comparable percent of CD4+ T cells producing IFN-γ, IL-17, and both cytokines (Figure 3a,b), indicating similar tissue levels of Th1 and Th17 cells. We also found comparable levels of foxp3+ induced Tregs between α4−/−→ RAG-1−/− and α4+/+→ RAG-1−/− mice (Figure 3c,d). Taken together, our data suggest that lack of α4 integrin does not affect in vitro polarization or in vivo distribution of Th1, Th17, or induced Tregs in colitic mice.

Levels of T helper type 1 (Th1) and T helper type 17 (Th17) cells and regulatory T cells (Tregs) in the colons of colitic mice. Representative (a) dot plots and (b) combined data from 7 to 16 individually analyzed animals and at least two independent experiments. Cells were initially gated on viable CD4+ T cells. (c) Representative staining and (d) combined data from five individually analyzed mice per group shows foxp3+ cells among viable CD4+ T cells. Lines on scatter graphs indicate mean values.
Homing of α4−/− and α4+/+ T cells to the GALT and the intestinal mucosa
Short-term homing experiments were performed next to compare the recruitment of α4−/− and α4+/+ T cells into the secondary lymphoid tissues and the intestinal mucosa. Homing of α4−/− T cells to the spleen, peripheral LN, and MLNs was largely unperturbed (Figure 4a, b), whereas homing to the intestine was notably reduced (Figure 4a,c). Phenotypic analysis of T cells recovered from these organs was consistent with the preferential homing of naive T cells for the secondary lymphoid tissues and activated/memory T cells to the extralymphoid tissues (Figure 4d). Taken together, these data suggest that lack of colitis in the α4−/−→RAG-1−/− mice is not likely due to the defective priming of these cells in the MLNs but is due to a defect in effector T-cell homing to the gut.

Overnight homing of activated/memory α4−/− T cells to the intestinal lamina propria (LP) is reduced. CD4+ T cells from spleens of α4−/− and α4+/+ mice were isolated, labeled with carboxyfluorescein succinimidyl ester (CFSE), and adoptively transferred into recipients. After an overnight homing (∼18 h), cells from the indicated tissues were isolated for flow cytometry analysis and homing index (HI) calculations were calculated as described in the Methods section. (a) Representative contour plots (initially gated on viable CD4+ cells) show percentages of α4−/− and α4+/+ T cells in the indicated tissues. Peripheral LNs (PLNs) were pooled from mandibular, cervical, and axillary LNs. (b, c) Scattergraphs show homing indices from three mice per group that were calculated based on the percentages shown in (a) and normalized to the input ratio of α4−/− and α4+/+ T cells. Line represents mean values. (d) Activation status of T cells isolated from the indicated organs after an overnight homing.
Contribution of T cell-associated α4 integrins during active colitis
Our experiments so far revealed a critical role of α4 integrin in T-cell homing to the noninflamed gut. The effectiveness of natalizumab in some CD patients, however, suggests that α4 may be also involved in homing to the inflamed intestines. Because α4−/−→RAG-1−/− mice do not develop colitis, we could not differentiate between contributions of T cell-associated α4 during the initiation of colitis from its role in established disease. To circumvent this, we cotransferred CD45.1 wild-type (WT) and CD45.2 α4−/− CD4+CD45RBhigh T cells into the same immunodeficient recipients. After disease developed, we compared the frequency of WT and mutant T cells in MLNs and colon LP of recipient mice. We found that the number of α4−/− T cells was significantly reduced compared with WT T cells (Figure 5a,b). This was not because of the reduced in vivo proliferation rates of α4−/− T cells as assessed by incorporation of bromodeoxyuridine (Supplementary Figure S7), thus suggesting that α4−/− T cells are defective in their recruitment into the inflamed colon. We also performed intracellular staining and evaluated production of IFN-γ and IL-17 by colon LP T cells. In agreement with data obtained from singly transferred α4−/− and α4+/+ CD4+CD45RBhigh T cells, we found comparable levels of IFN-γ+ cells among WT and α4−/− CD4 T cells in the cotransferred mice (Figure 5c,d). However, we detected fewer α4−/− CD4 T cells producing IL-17 and IFN-γ/IL-17 compared with WT T cells (Figure 5c,d). Levels of foxp3+ induced Tregs were similar among WT and α4−/− CD4 T cells (Figure 5e,f). These data suggest that α4 integrins are necessary for effector T-cell recruitment into MLNs and colon LP during active colitis. In addition, lack of this molecule does not affect tissue distribution of Th1 cells or Tregs but may reduce generation and/or accumulation of Th17 cells.

α4 Is important for T-cell localization to mesenteric lymph nodes (MLNs) and colon in colitic mice. Wild-type (WT; CD45.1) and α4−/− (CD45.2) CD4+CD45RBhigh T cells were coinjected (1:1 ratio, 0.5 × 106 each cell type per mouse) into recombination activating gene-1-deficient (RAG-1−/−) mice. After colitis developed (6–7 weeks), mice were killed and their MLN and colon lamina propria (LP) were dissected for T-cell analysis using flow cytometry. (a) T cells were initially gated on CD4+T-cell receptor (TCR)αβ+ cells. Representative contour plots from colon LP and MLN showing percent of WT and α4−/− T cells. (b) Absolute numbers of CD4+ T cells. For all bar graphs in this figure, combined data from five individually analyzed mice are expressed as mean±s.e.m. (c) After isolation, cells from colon LP and MLN were restimulated with phorbol 12-myristate 13-acetate (PMA)/ionomycin in the presence of GolgiStop for 4 h followed by intracellular staining to detect interferon-γ (IFN-γ) and interleukin-17 (IL-17). (c) Representative plots gated on WT or α4−/− TCRαβ+CD4+ T cells and (d) combined data are shown. (e) Representative contour plots and (f) pooled data show foxp3+ induced regulatory T cells (Tregs) among colon LP cells. Significance is indicated as*P<0.05 and ***P<0.001.
CD4+CD45RBhigh T cells lacking β1 integrin are capable of inducing chronic colitis
Prominent expression of LFA-1, ICAM-1, and β1 integrins on colon LP T cells suggested that these molecules might be important for T cells during colitis development. Our previous data showed that T cells lacking LFA-1 but not ICAM-1 are unable to trigger chronic colitis.28, 29 At the same time, the role of β1 integrins in colitis has not been assessed. Moreover, upregulation of VCAM-1 on endothelial cells in active CD22 suggests that circulating T cells can be recruited to the inflamed gut via α4β1/VCAM-1. To test the importance of β1 integrins in colitis development, we adoptively transferred CD4+CD45RBhigh T cells lacking β1 integrin from β1loxP/loxPxCD4-Cre+ (β1−/−) mice30 or β1-sufficient littermate control β1loxP/loxPxCD4-Creneg (β1+/+) mice into immunodeficient recipients and observed development of colitis in these mice. Absence of surface expression of β1 integrin on β1−/− T cells was confirmed by flow cytometry (Supplementary Figure S8A). Expression of other surface integrins was normal, including LPAM-1, that was in contrast to the initial findings with these mice.30 Following adoptive transfer into immunodeficient recipients, β1−/− T cells induced severe colitis with similar disease kinetics as did mice that received β1+/+ T cells (Figure 6a,b).

CD4+CD45RBhigh T cells lacking β1 integrins (β1−/−) produce severe colitis. (a) Blinded histopathological scores of colons from recipient immunodeficient recombination activating gene-1-deficient (RAG-1−/−) mice injected with β1+/+ (black circles) or β1−/− (white circles) T cells. (b) Representative hematoxylin and eosin (H&E)-stained images of distal colons were taken at × 100 magnification. (c) Absolute numbers of CD4+ T cell isolated from spleen, mesenteric lymph nodes (MLNs), colonic lamina propria (LP), and bone marrow (BM) of colitic mice. Pooled data from at least three independent experiments are shown. (d) Representative dot plots showing intracellular cytokine staining for interferon-γ (IFN-γ) and interleukin-17 (IL-17) by colon LP T cells isolated from colitic mice. Graphs shown were initially gated on viable CD4+ T cells. (e) Percentages of cells as shown in (d) pooled from 5 to 7 individually analyzed animals from at least two independent experiments. (f) Expression of integrin molecules on surface of CD4+ T cells isolated from colitic mice and analyzed using flow cytometry. Scatter plots show median fluorescent intensity (MFI) values. Shown are combined data with three animals from a representative experiment that was repeated twice. For all scatter plots, each circle represents an individual mouse; bars represent mean values per group. Significant differences between β1+/+ and β1−/− groups are indicated as*P<0.05 and ***P<0.001.
We next examined the effect of β1 integrin deletion on T-cell distribution in tissues. Absolute numbers of T cells in spleen, MLNs, colon LP, and bone marrow between mice that received β1+/+ and β1−/− T cells were comparable (Figure 6c). In addition, expression of α4 and LPAM-1 integrins on their surface were also similar (Supplementary Figure S6D). To determine whether lack of β1 integrins might also affect in vivo polarization of T cells into Th1, Th2, or Tregs, we analyzed percent of colon LP T cells positive for IFN-γ, IL-17, or foxp3 expression, respectively. We saw comparable percentages of IFN-γ+, IL-17+, and IFN-γ+IL-17+ T cells between the two groups (Figure 6d,e), indicating that lack of this molecule does not affect T-cell polarization in vivo. Levels of foxp3+ T cells in the colon LP T cells between the groups were also similar (Supplementary Figure S8B). Finally, expression of surface integrins on T cells isolated from different tissues from reconstituted mice were similar with the exception of β1 integrin that was, as expected, absent from the surface of β1−/− T cells (Figure 6f and Supplementary Figure S8C).
IBD patients treated with natalizumab show reduced accumulation of total CD4 T cells in the intestinal mucosa
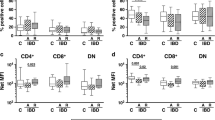
Although natalizumab has been approved for the treatment of CD, little is known about how this treatment affects distribution of pathogenic and Tregs in the intestinal mucosa of patients. We used immunohistochemistry to quantify total CD4+ and CD8+ T cells, Th1, and Tregs (Supplementary Figure S9) in biopsy samples taken from CD patients before, during, and after therapy with natalizumab. As patients were not consistently sampled at all three time points, longitudinally paired analyses were not possible, but by unpaired analyses, treatment with natalizumab significantly reduced the absolute number of CD4 but not CD8 T cells per tissue section area (Figure 7a). Tissue levels of Treg and Th1 CD4 T cells were not significantly affected, although a slight trend toward reduction was observed (Figure 7b). The ratio of Tregs and Th1 cells to total CD4 or CD8 T cells, or to one another, was not significantly altered with treatment (Figure 7c). Thus, the immunosuppressive effect of integrin α4 blockade on CD appears to result from a global reduction of CD4 T cells in the mucosa rather than a specific enrichment in the ratio of regulatory to pathogenic T cells.

Natalizumab decreases overall CD4 T-cell accumulation in the intestinal mucosa without differentially affecting T helper type 1 (Th1) cells or regulatory T cells (Tregs). (a) Intestinal biopsy specimens were processed for immunohistochemistry (IHC) and quantified as described in the Methods section. Comparison of CD4+ and CD8+ T cells between cohorts sampled before (pre), during (on), or after (post) natalizumab therapy were made by analysis of variance (ANOVA), using Tukey’s post-test. *P<0.05. (b) To quantify the CD4+ T-cell subsets per tissue section area, the number of individual Foxp3+ Tregs (left panel) and T-bet+ Th1 cells (right panel) were divided by the number of thousands of pixels staining positive for hematoxylin in their respective section, and comparisons between cohorts were made as in (a). (c) The ratio of Foxp3+ Tregs or Tbet+ Th1 cells to CD4+ or CD8+ pixels (each normalized to its section’s hematoxylin+ surface area) was calculated for each biopsy. Comparisons made as in (a) revealed no significant differences.
DISCUSSION
Prevention of immune cell infiltration with selective “anti-adhesion” therapies represents a promising strategy to treat IBD. Recent clinical studies showed that targeting α4 integrins with natalizumab leads to a significant improvement of disease parameters in patients with CD.2, 14, 31 Although this therapy was approved for the treatment of moderate-to-severe CD by Food and Drug Administration in 2008, the exact mechanism of action and the immune cells that are being targeted with this therapy have not been clearly defined. As effector T cells play an important role in maintaining chronic inflammation in IBD patients and experimental models,32, 33 natalizumab is thought to work by blocking circulating T cells from entering the inflamed intestine. However, this idea has been largely supported by indirect evidence. Because α4β7 is expressed on several cell types, including B cells, natural killer cells, monocytes, and eosinophils, it is not clear whether the beneficial effects of the different antibody preparations is directed against one or more of these leukocyte populations. Therefore, we addressed this question directly using adoptive transfer of T cells lacking surface expression of α4 integrin.
We found that adoptive transfer of α4neg or α4−/− T cells produced significantly attenuated colitis in RAG-1−/− mice compared with those with intact α4 integrin. In agreement with previous studies,26, 27 our data show that lack of α4 expression does not affect activation, proliferation, or polarization of naive T cells to a pathogenic Th1/Th17 phenotype in vitro or in vivo. Our short-term homing experiments revealed that migration of naive T cells to the MLNs was similar between α4−/− and α4+/+ T cells. However, recruitment of activated/memory/effector T cells lacking surface α4 to the small intestine or colon LP was significantly compromised. These data are in agreement with the existing paradigm34, 35 that under normal conditions, homing to the GALT involves redundant use of L-selectin and α4β7, whereas recruitment to small and large intestine is largely dependent on the latter molecule.
Which surface molecules are important for the homing of Th1 and Th17 cells is a subject of intense debate. As both subsets have been implicated in the pathogenesis of IBD, it is critical to delineate mechanisms how these cells traffic to the gut. Using β7−/− mice, Wang et al. 35 demonstrated a critical role of α4β7 for in vitro-polarized Th17 cell migration to the entire intestine. However, compared with Th1 cells, Th17 represent a relatively small population of LP T cells in colitic mice (Figure 3b). In agreement with previous reports,26, 27 we found that expression of α4 integrin is substantially higher on the surface of Th1 cells compared with Th17 cells. Although loss of α4 integrin led to fewer Th1 cells and more Th17 cells accumulating in the central nervous system during experimental autoimmune encephalomyelitis development,26, 27 accumulation of Th1 cells or Th1/Th17 ratio in the gut was not perturbed even under the identical inflammatory environment that was created in mice cotransferred with WT and α4−/− T cells. These findings in mice were corroborated by data obtained in human CD patients. Treatment with natalizumab led to a global reduction in intestinal CD4 T cells but did not affect Th1 or Tregs. Whether this correlates with sequestration of effector T cells in peripheral blood, as recently reported in natalizumab-treated multiple sclerosis patients,36 or not remains to be established. Taken together, our data suggest that deficiency in α4 broadly affects CD4 T-cell trafficking to the gut without preferentially affecting one Th subset more than the other.
Analysis of α4 and α4β7 integrin expression on the surface of T cells in colitic mice revealed dynamic tissue-specific changes between spleen, MLNs, and colonic LP (Figure 2a). Whereas T cells isolated from MLNs expressed α4β7, it was not detected on those cells isolated from colon. Indeed, decreased expression of α4β7 by intestinal T lymphocytes has been reported in IBD patients.37 Thus, our data suggest that following migration into the intestine, T cells lose expression of gut-homing molecules α4β7 and CCR9 (data not shown). It is not clear at the present time if this occurs only under inflammatory conditions or may also occur following migration of T cells into noninflamed intestine.
In contrast to the lack of α4β7, β1 integrin was highly expressed on the surface of colon LP T cells in colitic mice. The β1 integrin subunit can form heterodimers with α1–α6 and αV α-chains to form a family of very late antigen molecules that can bind to extracellular matrix proteins laminin, fibronectin, collagen, and others.38 Although expression of β1 integrin may be important for T cells to receive survival and/or additional costimulatory signals in colon or bone marrow to maintain their effector functions during colitis, adoptive transfer of T cell lacking this integrin did not protect mice from developing colitis. We also found that T-cell numbers in the colon, secondary lymphoid tissues, and bone marrow were comparable. Therefore, among the α4 integrins, α4β7 plays a critical and nonredundant role in colitis pathogenesis, whereas α4β1 appears to be dispensable for the colitis-inducing abilities of T cells.
Taken together, our study demonstrated a critical role for T cell-associated α4 integrins in experimental colitis and human IBD. We directly assessed the relative importance of T cell-associated α4β7 and α4β1 in the induction of chronic, T cell-mediated colitis. In addition to the role of α4 integrins in induction of colitis, we definitively showed importance of these molecules for T cell’s ability to traffic to MLNs and colon during active disease. Furthermore, our data with natalizumab-treated CD patients corroborated our findings in mice, by showing, for the first time, that natalizumab treatment reduced accumulation of CD4 T cells in the inflamed intestines. Finally, our results revealed an interesting dynamic of gut-homing molecule expression between circulating and tissue-associated T cells. Better understanding of the mechanisms that regulate surface integrin expression in effector T cells during their intravascular trafficking and beyond might provide additional clues to designing novel anti-adhesion therapies for IBD and related conditions.
METHODS
Animals. WT mice and RAG-1−/− mice (B6.129S7-Rag1tm1Mom/J) on the C57Bl/6 background were purchased from The Jackson Laboratory (Bar Harbor, ME). α4loxP/loxP Mice on a C57Bl/6 background were kindly provided by Dr Thalia Pappayanapoulou (University of Washington, Seattle, WA).39 The β1loxP/loxPxCD4-Cre+ (β1−/−) mice were kindly provided by Dr Yoji Shimizu (University of Minnesota, Minneapolis, MN). Mx.cre and CD4.Cre mice were purchased from the Taconic Farms (Madison, CT). To generate α4-deficient T cells, α4loxP/loxP x Mx.cre+ mice were injected with 300–450 μg of polyI:C intraperitoneally in 1 × phosphate-buffered saline every other day for a total of seven injections. At 2 weeks after the last injection, mice were bled from the tail vein to determine the efficiency of α4 deletion from the surface of T cells using flow cytometry. Mice with <20% α4-negative cells of the total CD4+ T cells were given additional 3–5 polyI:C injections. All mice that were used in subsequent experiments were used at least 2 weeks after the last polyI:C injection. To generate mice with specific ablation of α4 or β1 in T cells, we crossed α4loxP/loxP or β1loxP/loxP mice with CD4.Cre mice.40 Animals were maintained on 12 h/12 h light/dark cycles in standard animal cages with filter tops under specific pathogen-free conditions at the animal care facility of the Louisiana State University Health Sciences Center and given standard laboratory rodent chow and water ad libitum. All experimental procedures involving the use of animals were reviewed and approved by the Institutional Animal Care and Use Committee of Louisiana State University Health Sciences Center in Shreveport and performed according to the criteria outlined by the National Institute of Health.
Antibodies. CD62L (L-selectin)-fluorescein isothiocyanate (FITC), CD44-phycoerythrin (PE), CD25-PE, CD3-PE-Cy5, and CD4-allophycocyanin were used for quantifying T-cell numbers in different tissues; CD11a-FITC, CD49d-PE, CD103-FITC, β7 integrin-peridinin chlorophyll protein-Cy5.5 (PerCP-Cy5.5), ICAM-1-PE, CD18-FITC, CD29-PE, and CD62L-FITC were used for analysis of surface expression of T-cell adhesion molecules. Antibodies were obtained from either BD Biosciences (San Diego, CA) or eBioscience (San Diego, CA).
Adoptive T-cell transfer model of colitis. For adoptive T-cell transfer experiments, spleens from several α4loxP/loxP Mx.cre+ mice or polyI:C-injected α4loxP/loxP Mx.cre- mice were removed and pooled. Splenocytes were prepared for sorting as previously described.41 Cells were stained with anti-CD4, -CD45RB, and -CD49d antibodies. For sorting CD4+CD45RBhigh cells (CD4+ cells with 40% of the brightest expression of the CD45RB) were sorted into CD49d-negative (CD49dneg) and CD49d+ populations using FACSAria (Becton-Dickinson, San Jose, CA) and were found to be >95% pure on post-sort analysis. Age- and sex-matched RAG-1−/− mice were injected intraperitoneally with 5 × 105 CD4+CD45RBhighCD49d+ or CD4+CD45RBhighCD49dneg T cells resuspended in 500 μl of phosphate-buffered saline. Control groups included RAG-1−/− mice injected with 5 × 105 of the CD4+CD45RBhigh obtained either from WT mice or from Mx.cre-mice treated with polyI:C at least 7 times (300–450 μg per mouse, intraperitoneally every other day).
For α4loxP/loxPCD4-cre+ (α4−/−), α4loxP/loxPCD4-creneg (α4+/+), β1loxP/loxPCD4-cre+ (β1−/−), and β1loxP/loxPCD4-creneg (β1+/+) adoptive transfers, spleens from several mice were pooled and purified by fluorescence-activated cell sorting into CD4+CD45RBhigh cells, and injected into age- and sex-matched RAG-1−/− mice. Clinical evidence of disease (e.g., body weight loss and loose stool/diarrhea) was followed and recorded weekly from the time of the injection. At 8 weeks after transfer or when the animals lost 15–20% of their original body weights, mice were killed and their colons were excised and processed for histopathology and blinded scoring using previously published criteria.41 For coinjection studies, WT (CD45.1) and α4−/− (CD45.2) CD4+CD45RBhigh T cells were coinjected (1:1 ratio, 0.5 × 106 each cell type per mouse) into RAG-1−/− mice.
Mononuclear cell isolation and analyses. Mononuclear cells were isolated from the spleen, MLNs, and colonic LP of reconstituted recipient mice and analyzed using flow cytometry as previously described.28, 41, 42 For intracellular staining, mononuclear cells were activated overnight with antibodies to CD3 and CD28 with GolgiStop (BD Biosciences) added for the last 6 h before processing for surface staining, fixation, and permeabilization, followed by intracellular staining using antibodies and kits from eBioscience according to the manufacturer’s protocol.
Cytokine measurement. Mononuclear cells were cultured in 96-well plate (100,000/well) in the presence of activating anti-CD3/CD28 antibodies for 24 h. Cytokine concentrations in culture supernatants were measured using 22-plex Mouse Cytokine/Chemokine Immunoassay (Millipore, Billerica, MA). For serum cytokine determinations, blood was collected from reconstituted mice, and serum was separated, aliquoted, and kept frozen until it was analyzed using mouse 32-plex (Millipore) according to the manufacturer’s instructions. Samples were run on a Bio-Plex Luminex instrument (Bio-Rad Laboratories, Hercules, CA). Only those values that fell within the range of the standard curve were used in calculations unless otherwise specified. Data were analyzed using Miliplex Analyzer 3.1 xPONENT software (Millipore). For the cytokines that were out of range, cytokine-specific enzyme-linked immunosorbent assay was used (eBioscience) according to the manufacturers’ protocol.
T-cell homing studies. For homing experiments, CD4+ T cells from α4−/− and α4+/+ control mice were isolated from spleens using EasySep CD4+ negative selection kit (StemCell Technologies, Vancouver, BC, Canada) according to the manufacturer’s protocol. Cells were labeled with carboxyfluorescein succinimidyl ester (Life Technologies) or CellTracker Blue CMAC (7-Amino-4-Chloromethylcoumarin) (Life Technologies, Carlsbad, CA) according to the manufacturer’s protocol. Cells were washed, resuspended at 10 × 107/ml in phosphate-buffered saline, and 2 × 107 cells (in 100–200 μl) were injected retro-orbitally into anesthetized RAG-1−/− mice. At different times, mice were killed and the indicated tissues were harvested and processed as described above. For analysis, ∼1–2 × 106 of FcR-blocked cells were placed in individual wells of a 96-well plate and stained with fluorescently labeled antibodies to CD4. Mice were killed at indicated times and homing index of α4+/+ and α4−/− cells in different tissues (HITISSUE) was calculated according to the method by Villablanca et al.43 using the following formula:

Human studies. Through an institutional review board-approved protocol, electronic medical records were reviewed at a single hospital (Virginia Mason Medical Center, Seattle, WA) to identify 10 patients with CD (age 25–60, median 38, 50% female) who underwent colonoscopic biopsies before, during (e.g., within 1 month of last dose), or after (>1.5 months since last dose) treatment with natalizumab. Five patients were also on infliximab, four were on a thiopurine, and eight were on a glucocorticoid during one or more biopsies. Paraffin blocks of surplus formalin-fixed tissue from these biopsies were then retrieved from the hospital’s pathology archive, and sectioned in a de-identified manner. Limited clinical data on the patients biopsied were similarly retrieved and stored in a secure and de-identified manner. All work was performed in compliance with human subject research ethical standards and medical privacy laws.
Immunohistochemistry for human patients with CD. Tissue sections were stained by immunohistochemistry with antibodies to either FOXP3 (clone 236A/E7, eBioscience, San Diego, CA), CD4 (clone 4B12, Vector Labs, Burlingame, CA), or a combination of T-bet (clone 4B10, Santa Cruz Biotech, Santa Cruz, CA) and CD8 (clone 4B11, Novocastra, Devil, FL). Sections were photographed digitally in their entirety with the TissueFAXS platform (Tissue Gnostics, Tarzana, CA).
Because CD4+ and CD8+ cells were often too confluent to reliably identify individual positive cells, the number of pixels staining positive for CD4 (Figure 7a, left panel) or CD8 (Figure 7a, right panel) were divided by the number of pixels staining positive for hematoxylin in their respective section. To quantify the CD4+ T-cell subsets per tissue section area, the number of individual Foxp3+ Tregs (Figure 7b, left panel) and T-bet+ Th1 cells (Figure 7b, right panel) were divided by the number of thousands of pixels staining positive for hematoxylin in their respective section, and comparisons between cohorts were made as in Figure 7a.
Staining for CD4 was not strong enough for two-color immunohistochemistry, but all detectable T-bet was detected in CD3+ T cells (data not shown), virtually all of which in the LP are known to be either CD8+ or CD4+. Therefore, to quantify Th1 cells (which should be CD4+, T-bet+), T-bet+/CD8− cells were manually counted within digital images by a blinded observer (J.D.L.) Foxp3+ cells in the LP (all of which are CD4+ by flow cytometry, data not shown) were counted digitally with ImageJ software (NIH, Bethesda, MD), using a color deconvolution program to isolate positively stained pixels. Because of confluency in certain areas, CD4+ and CD8+ cells could not be manually counted, and hence to measure the relative density of these cells in tissue section images, the number of pixels staining positive for either marker was quantified with ImageJ, and normalized against cells staining with hematoxylin (a nonspecific nuclear stain) to control for variations in total cellularity in each section.
Statistics. Statistical analyses were done using GraphPad Prism software (version 5, La Jolla, CA). Unless otherwise indicated, data are presented as mean±s.e.m. Statistical significance between any two groups was evaluated using unpaired two-sided t-tests. Statistical significance between more than two groups was evaluated using a one-way analysis of variance. Statistical significance between selected groups was evaluated using Dunnett’s multiple comparison test. A probability (P-value) of <0.05 was considered significant.
References
Feagan, B.G. et al. Treatment of active Crohn’s disease with MLN0002, a humanized antibody to the alpha4beta7 integrin. Clin. Gastroenterol. Hepatol. 6, 1370–1377 (2008).
Ghosh, S. et al. Natalizumab for active Crohn's disease. N. Engl. J. Med. 348, 24–32 (2003).
Park, E.J. et al. Aberrant activation of integrin alpha4beta7 suppresses lymphocyte migration to the gut. J. Clin. Invest. 117, 2526–2538 (2007).
Podolsky, D.K. et al. Attenuation of colitis in the cotton-top tamarin by anti-alpha 4 integrin monoclonal antibody. J. Clin. Invest. 92, 372–380 (1993).
Podolsky, D.K. Selective adhesion-molecule therapy and inflammatory bowel disease—a tale of Janus? N. Engl. J. Med. 353, 1965–1968 (2005).
Rivera-Nieves, J. et al. L-selectin, alpha 4 beta 1, and alpha 4 beta 7 integrins participate in CD4+ T cell recruitment to chronically inflamed small intestine. J. Immunol. 174, 2343–2352 (2005).
Yang, J.T., Rayburn, H. & Hynes, R.O. Cell adhesion events mediated by alpha 4 integrins are essential in placental and cardiac development. Development 121, 549–560 (1995).
Sydora, B.C. et al. beta7 Integrin expression is not required for the localization of T cells to the intestine and colitis pathogenesis. Clin. Exp. Immunol. 129, 35–42 (2002).
Kurmaeva, E, Boktor, M, Zhang, S, Bao, R, Berney, S. & Ostanin, D.V. Roles of T cell-associated L-selectin and beta7 integrins during induction and regulation of chronic colitis. Inflamm. Bowel Dis. 19, 2547–2559 (2013).
Picarella, D, Hurlbut, P, Rottman, J, Shi, X, Butcher, E. & Ringler, D.J. Monoclonal antibodies specific for beta 7 integrin and mucosal addressin cell adhesion molecule-1 (MAdCAM-1) reduce inflammation in the colon of scid mice reconstituted with CD45RBhigh CD4+ T cells. J. Immunol. 158, 2099–2106 (1997).
Bjursten, M, Bland, P.W., Willen, R. & Hornquist, E.H. Long-term treatment with anti-alpha 4 integrin antibodies aggravates colitis in G alpha i2-deficient mice. Eur. J. Immunol. 35, 2274–2283 (2005).
Hesterberg, P.E. et al. Rapid resolution of chronic colitis in the cotton-top tamarin with an antibody to a gut-homing integrin alpha 4 beta 7. Gastroenterology 111, 1373–1380 (1996).
Targan, S.R. et al. Natalizumab for the treatment of active Crohn’s disease: results of the ENCORE Trial. Gastroenterology 132, 1672–1683 (2007).
Sandborn, W.J. et al. Natalizumab induction and maintenance therapy for Crohn’s disease. N. Engl. J. Med. 353, 1912–1925 (2005).
Feagan, B.G. et al. Vedolizumab as induction and maintenance therapy for ulcerative colitis. N. Engl. J. Med. 369, 699–710 (2013).
Sandborn, W.J. et al. Vedolizumab as induction and maintenance therapy for Crohn’s disease. N. Engl. J. Med. 369, 711–721 (2013).
Hirahara, K, Liu, L, Clark, R.A., Yamanaka, K, Fuhlbrigge, R.C. & Kupper, T.S. The majority of human peripheral blood CD4+CD25highFoxp3+ regulatory T cells bear functional skin-homing receptors. J. Immunol. 177, 4488–4494 (2006).
Lord, J.D., Valliant-Saunders, K, Hahn, H, Thirlby, R.C. & Ziegler, S.F. Paradoxically increased FOXP3+ T cells in IBD do not preferentially express the isoform of FOXP3 lacking exon 2. Dig. Dis. Sci. 57, 2846–2855 (2012).
Bowden, R.A. et al. Role of alpha4 integrin and VCAM-1 in CD18-independent neutrophil migration across mouse cardiac endothelium. Circ. Res. 90, 562–569 (2002).
Schneider, M.K., Strasser, M, Gilli, U.O., Kocher, M, Moser, R. & Seebach, J.D. Rolling adhesion of human NK cells to porcine endothelial cells mainly relies on CD49d-CD106 interactions. Transplantation 73, 789–796 (2002).
Soler, D, Chapman, T, Yang, L.L., Wyant, T, Egan, R. & Fedyk, E.R. The binding specificity and selective antagonism of vedolizumab, an anti-α4β7 integrin therapeutic antibody in development for inflammatory bowel diseases. J. Pharm. Exp. Ther. 330, 864–875 (2009).
Kawachi, S. et al. Cytokine and adhesion molecule expression in SCID mice reconstituted with CD4+ T cells. Inflamm. Bowel Dis. 6, 171–180 (2000).
Berlin, C. et al. Alpha 4 beta 7 integrin mediates lymphocyte binding to the mucosal vascular addressin MAdCAM-1. Cell 74, 185–195 (1993).
Strauch, U.G., Lifka, A, Gosslar, U, Kilshaw, P.J., Clements, J. & Holzmann, B. Distinct binding specificities of integrins alpha 4 beta 7 (LPAM-1), alpha 4 beta 1 (VLA-4), and alpha IEL beta 7. Int. Immunol. 6, 263–275 (1994).
Wagner, N. et al. Critical role for beta7 integrins in formation of the gut-associated lymphoid tissue. Nature 382, 366–370 (1996).
Rothhammer, V. et al. Th17 lymphocytes traffic to the central nervous system independently of {alpha}4 integrin expression during EAE. J. Exp. Med. 208, 2465–2476 (2011).
Glatigny, S, Duhen, R, Oukka, M. & Bettelli, E. Cutting edge: loss of alpha4 Integrin expression differentially affects the homing of Th1 and Th17 cells. J. Immunol. 187, 6176–6179 (2011).
Ostanin, D.V. et al. T cell-associated CD18 but not CD62L, ICAM-1, or PSGL-1 is required for the induction of chronic colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 292, G1706–G1714 (2007).
Pavlick, K.P. et al. Role of T-cell-associated lymphocyte function-associated antigen-1 in the pathogenesis of experimental colitis. Int. Immunol. 18, 389–398 (2006).
DeNucci, C.C., Pagan, A.J., Mitchell, J.S. & Shimizu, Y. Control of alpha4beta7 integrin expression and CD4 T cell homing by the beta1 integrin subunit. J. Immunol. 184, 2458–2467 (2010).
Gordon, F.H. et al. A randomized placebo-controlled trial of a humanized monoclonal antibody to alpha4 integrin in active Crohn’s disease. Gastroenterology 121, 268–274 (2001).
Abraham, C. & Cho, J.H. Inflammatory bowel disease. N. Engl. J. Med. 361, 2066–2078 (2009).
Sartor, R.B. Mechanisms of disease: pathogenesis of Crohn’s disease and ulcerative colitis. Nat. Clin. Pract. Gastroenterol. Hepatol. 3, 390–407 (2006).
Hamann, A, Andrew, D.P., Jablonski-Westrich, D, Holzmann, B. & Butcher, E.C. Role of alpha 4-integrins in lymphocyte homing to mucosal tissues in vivo. J. Immunol. 152, 3282–3293 (1994).
Wang, C, Kang, S.G., HogenEsch, H, Love, P.E. & Kim, C.H. Retinoic acid determines the precise tissue tropism of inflammatory Th17 cells in the intestine. J. Immunol. 184, 5519–5526 (2010).
Kivisakk, P. et al. Natalizumab treatment is associated with peripheral sequestration of proinflammatory T cells. Neurology 72, 1922–1930 (2009).
Meenan, J, Spaans, J, Grool, T.A., Pals, S.T., Tytgat, G.N. & van Deventer, S.J. Altered expression of alpha 4 beta 7, a gut homing integrin, by circulating and mucosal T cells in colonic mucosal inflammation. Gut 40, 241–246 (1997).
Springer, T.A. Adhesion receptors of the immune system. Nature 346, 425–434 (1990).
Scott, L.M., Priestley, G.V. & Papayannopoulou, T. Deletion of alpha4 integrins from adult hematopoietic cells reveals roles in homeostasis, regeneration, and homing. Mol. Cell. Biol. 23, 9349–9360 (2003).
Lee, P.P. et al. A critical role for Dnmt1 and DNA methylation in T cell development, function, and survival. Immunity 15, 763–774 (2001).
Ostanin, D.V. et al. T cell transfer model of chronic colitis: concepts, considerations, and tricks of the trade. Am. J. Physiol. Gastrointest. Liver Physiol. 296, G135–G146 (2009).
Ostanin, D.V. et al. T cell-induced inflammation of the small and large intestine in immunodeficient mice. Am. J. Physiol. Gastrointest. Liver Physiol. 290, G109–G119 (2006).
Villablanca, E.J. & Mora, J.R. Competitive homing assays to study gut-tropic t cell migration. J. Vis. Exp. 29, G2619 (2011).
Acknowledgements
D.V.O. is supported by the Center of Excellence for Arthritis and Rheumatology funds and CCFA Career Developmental Award 2923. M.B.G. is supported by NIH/NIDDK (RO1-091269). J.D.L. is supported by NIH/NIDDK (K08 DK081659) and NIH/NIAID (U19 AI050864). We sincerely thank Drs Thalia Pappayanapoulou and Tatiana Ulyanova (University of Washington) for providing α4loxP/loxP mice and for their assistance in genotyping, and Drs Yoji Shimizu and Brandon Burbach for providing β1loxP/loxP mice. We are thankful to Dr Robert Chervenak, Deborah Chervenak, and Shannon Mumphrey for their technical expertise and assistance with flow cytometry and to Talicia Tarver for critical reviewing of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declared no conflict of interest.
Additional information
SUPPLEMENTARY MATERIAL is linked to the online version of the paper
Supplementary information
Rights and permissions
About this article
Cite this article
Kurmaeva, E., Lord, J., Zhang, S. et al. T cell-associated α4β7 but not α4β1 integrin is required for the induction and perpetuation of chronic colitis. Mucosal Immunol 7, 1354–1365 (2014). https://doi.org/10.1038/mi.2014.22
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mi.2014.22
This article is cited by
-
IL-36 cytokines imprint a colitogenic phenotype on CD4+ T helper cells
Mucosal Immunology (2022)
-
Dual functions of Rap1 are crucial for T-cell homeostasis and prevention of spontaneous colitis
Nature Communications (2015)
