
Abstract
Vanilloid receptor 1 (VR1) is expressed on immune cells as well as on sensory neurons. Here we report that VR1 can regulate immunological events in the gut in response to its ligand Capsaicin (CP), a nutritional factor, the pungent component of chili peppers. Oral administration of CP attenuates the proliferation and activation of autoreactive T cells in pancreatic lymph nodes (PLNs) but not other lymph nodes, and protects mice from development of type 1 diabetes (T1D). This is a general phenomenon and not restricted to one particular strain of mice. Engagement of VR1 enhances a discreet population of CD11b+/F4/80+ macrophages in PLN, which express anti-inflammatory factors interleukin (IL)-10 and PD-L1. This population is essential for CP-mediated attenuation of T-cell proliferation in an IL-10-dependent manner. Lack of VR1 expression fails to inhibit proliferation of autoreactive T cells, which is partially reversed in (VR1+/+ → VR1−/−) bone marrow chimeric mice, implying the role of VR1 in crosstalk between neuronal and immunological responses in vivo. These findings imply that endogenous ligands of VR1 can have profound effect on gut-mediated immune tolerance and autoimmunity by influencing the nutrient–immune interactions.
Similar content being viewed by others

Introduction
Vanilloid receptor 1 (VR1) is a transient receptor potential ion channel and is activated by various ligands such as Capsaicin (CP), protons, heat, and lipid metabolites.1, 2, 3 It is highly expressed on sensory neurons of dorsal root and trigeminal ganglia and on cells of the immune system, including dendritic cells (DCs) and macrophages.4, 5, 6 CP, a ligand for VR1, is the principal pungent component of red peppers. Chemically, it is a derivative of vanillyl amide, 8-methyl-N-vanillyl-6-noneamide. The engagement of VR1 on DCs by its ligand CP modulates their function by upregulating antigen-presenting and costimulatory molecules, in addition to initiating their migration to draining lymph nodes.4 The robust adjuvanticity of CP initiated the studies on its effect in tumor therapy and clearly demonstrated that intra-tumoral administration of CP leads to the regression of advanced pre-existing tumors, and such immunity is T-cell mediated and tumor-specific.7, 8 Considering that CP exhibits powerful immune consequences and is a natural product widely ingested as a food additive, we investigated if CP is immuno-modulatory when administered by oral route.
The gut-associated lymphoid tissue is exposed to myriads of antigens daily and must discern between harmless food antigens and commensal microorganisms from pathogen-derived antigen in the gastrointestinal tract. Antigen-presenting cells (APCs), including DCs and macrophages, have an essential role in maintaining the balance between tolerance and protective immunity in the gut.9 This balance is central to maintaining immune homeostasis. Mucosal DCs and macrophages have the ability to induce T-cell differentiation (Th1, Th2, Th17).10, 11 Moreover, both these populations exhibit regulatory function and have been described to induce T-regulatory cells (Tregs) in the gut.12, 13 APCs resident in the lamina propia, Peyer's patches, and mesenteric lymph nodes have direct access to intestinal bacteria, and antigens delivered by oral route are processed and presented by Peyer's patch DCs.14, 15 Interestingly, studies have shown that intestinal contents have significance in the pancreatic lymph nodes (PLNs). Specifically, fed antigens are presented in the PLN along with pancreatic antigens and have consequences for autoimmune destruction of the pancreas.16 In this manner the PLN is at the crux of the gastrointestinal tract and the pancreas, and can sample the environments of both, which has significance in type 1 diabetes (T1D).17, 18, 19
T1D is one of the most prevalent autoimmune diseases worldwide. It results from the destruction of the insulin-producing β-cells of the pancreas. The disease is mediated by T cells and results from a poorly understood breakdown of immune tolerance to self-antigens. Our understanding of the pathogenesis and etiology of T1D is incomplete, but the non-obese diabetic (NOD) mouse model of T1D is a well-studied model that shares many pathological and immunological features of human T1D. Many studies have evaluated the oral administration of immune modulators in the treatment of T1D in NOD.20, 21 Oral therapies hold great promise in treatment of T1D.
Here we examined the immune-modulatory properties of CP when administered by oral route in two mouse models of T1D. The potent influence of CP on the immune system is clear; however, our understanding of the required mode of delivery and the mechanisms by which the sensory receptor VR1 modulates an immune response is incomplete, in particular the role of VR1 expressing APCs in autoimmunity. Although engagement of VR1 has previously been shown to be proinflammatory, here we show the immunoregulatory influence of orally administered CP acting directly on the immune system in the context of T1D.
Results
Oral administration of CP inhibits antigen-specific T cells in PLN
VR1 ligand CP is a widely ingested food additive and has powerful immune consequences; therefore, we investigated the immuno-modulatory role of VR1 in the gut by orally administering CP. To test this we used two different mouse models, i.e., NOD and rat insulin promoter (RIP)-ovalbumin (OVA). NOD mice spontaneously develop insulitis by destruction of pancreatic β-cells similar to T1D in humans. Both CD4 and CD8 subsets of T cells are thought to be involved in the spontaneous development of insulitis and β-cell destruction in the NOD mouse. Moreover, the pancreas-draining lymph node (PLN) is considered to be a part of the gut-associated lymphoid tissue and the gastrointestinal tract, providing a conduit for environmental agents to directly modify the immune response to pancreatic autoantigens.16 Thus, we tested if orally administered CP has any consequence on β-cell antigen–specific T-cell proliferation in PLN. NOD mice were fed with vehicle or 10 μg of CP, and 24 h later received adoptive transfer of carboxyfluorescein succinimidyl ester (CFSE)-labeled BDC2.5 CD4+ T cells. BDC2.5 T-cell antigen receptor (TCR) transgenic (Tg) cells are specific for a pancreatic islet antigen. The proliferation of adoptively transferred BDC2.5 T cells was assessed 4 days post-transfer and showed a significant decrease in the T-cell proliferation in mice fed with 10 μg of CP in comparison with the vehicle control fed mice (Figure 1a).

Oral administration of Capsaicin (CP) attenuates antigen-specific T cells in the pancreatic lymph nodes (PLNs). (a) Representative fluorescence-activated cell sorting (FACS) analysis of prediabetic recipient non-obese diabetic (NOD) mice (n=3 per group) fed with CP (10 μg) or vehicle (veh) control and 24 h later received adoptive transfer intravenous of CFSE-labeled NOD.BDC2.5 CD4+CD25− cells (diabetogenic T-cell clone). At 3 days post-adoptive transfer, the PLNs were harvested and CFSE dilution was analyzed by FACS. Graph indicates average % (±s.d.) of BDC2.5 CD4+ cells that diluted CFSE 3 days post-transfer, out of total transferred BDC2.5 cells. Data are representative of three independent experiments. (b and c) Same experimental design as in panel a, except recipient mice were rat insulin promoter (RIP)-ovalbumin (OVA) mice (n=3 per group) that received CFSE-labeled CD45.1+CD8+ OT-I cell adoptive transfer. Data are representative of three independent experiments. (b) Representative histograms of RIP-OVA PLN CFSE dilution (n=3–4 per group; left panel). Graph indicates average % (±s.d.) of divided OT-I cells out of the total OT-I transferred population in the PLN (middle panel) and RLN (right panel). (c) Representative FACS analysis of CD62L and CD44 expression on CD45.1+CD8+ OT-I, numbers in quadrants indicate percent cells in each (left panel). Graph indicates average % CD62 L+CD44− (±s.d.) of OT-I cells out of total OT-I transferred population in the PLN (n=4 per group; right panel). (d) RIP-OVA-fed CP (10 μg) or vehicle control and 24 h later received transfer of CD45.1+CD8+ OT-I and PLNs were harvested after 5 days and restimulated in vitro with PMA + I in the presence of BFA for 4 h. Intracellular cytokine staining was performed for the cytokines indicated. Data are representative of two independent experiments. *P=0.05, **P=0.01. BFA, Brefeldin A; IFN, interferon; NS, nonsignificant; TNF, tumor necrosis factor.
We further confirmed these results using CD8+ T cells in an additional model with known pancreatic antigen, the RIP-OVA model. In RIP-OVA Tg mice, the model antigen OVA is expressed as a transgene under the control of the RIP and OVA is expressed as a self-antigen in the insulin-producing β-cells in the islets of Langerhans.22 Transfer of CFSE-labeled OT-I cells into RIP-OVA mice fed 24 h prior with 10 μg or 25 μg of CP or vehicle yielded similar results. OT-I T-cell proliferation was abrogated in mice that receive the 10 μg CP dose (Figure 1b, left and middle panel; Supplementary Figure S1a and b online). One possible explanation for the abrogation of proliferation is that the inhibitory dose of CP is toxic to T cells. To address this possibility, we compared the frequency of OT-I T cells in the PLN of C57BL/6 mice that were fed 10 μg of CP, or vehicle, and found no difference in OT-I survival after feeding (Supplementary Figure S2 online). As expected based on the CFSE profiles, we also observed an increase in the naive population (CD44lowCD62Lhigh) of T cells (Figure 1c) and reduced expression of effector cytokines (interferon-γ and tumor necrosis factor-α) in CP 10 μg fed mice (Figure 1d and Supplementary Figure S1c online). Therefore, these results indicate that orally administered CP can tolerize autoreactive CD4+ and CD8+ T cells in two distinct mice models, the NOD and RIP-OVA mice.
Inhibition of T-cell proliferation is localized to the gut
Considering that intestinal contents (antigens) are presented in the PLN,16 we were interested if the abrogation of T-cell proliferation was specific to the PLN, a gut-associated lymph node, or if feeding CP results in systemic suppression. We took advantage of one feature observed in RIP-OVA mice, where OVA is expressed in the proximal tubular cells of the kidney in addition to the pancreas.22 As adoptively transferred OT-I cells will proliferate in the renal lymph nodes (RLNs), we harvested RLN from RIP-OVA that had been fed and transferred CFSE-labeled OT-I. The results in Figure 1b (right panel) and Supplementary Figure S1a and b online indicate no difference in the proliferation of OT-I in the RLN after feeding vehicle, CP 10 μg, or 25 μg, in stark contrast to the proliferation profile observed in the PLN, where T-cell suppression is observed after the 10 μg dose. These results indicate that the suppression of T-cell proliferation after CP feeding is not systemic but localized to gut-draining lymph nodes.
Oral administration of CP inhibits spontaneous diabetes in NOD mice
NOD mice develop insulitis spontaneously, starting at about 5 weeks of age and clinical diabetes starting at 10–12 weeks of age. T1D is a result of destruction of the insulin-producing β-cells in the pancreas by autoreactive T cells. In order to determine whether CP-mediated inhibition of autoreactive T-cell proliferation and function can influence onset of T1D in NOD mice, we fed these mice (5 mice per group) with different doses of CP (0.1, 1, 10, 25, or 50 μg) on the 9th and 10th week of age. Control mice were either left untreated or fed with vehicle. Overall, 80% of untreated mice were diabetic at 17 weeks of age, whereas 60% of the mice fed with vehicle were diabetic at 15 weeks of age (Figure 2a–c). In contrast, 80–100% of mice fed with 10 μg CP remained disease free for 20 weeks. Lower doses of CP (i.e., 0.1 μg or 1 μg) and higher doses (i.e., 25 μg or 50 μg) did not show any protection (Figure 2a–c). Histological examination (representative of 14 different islets from four CP 10 μg fed mice) revealed a correlation between urine glucose levels and lymphocytic islet infiltration in the pancreas (Figure 2d). We next turned our attention to identify the cell population responsible for the inhibition of autoreactive T-cell proliferation.

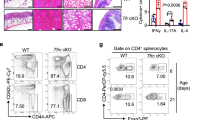
Oral administration of Capsaicin (CP) protects non-obese diabetic (NOD) mice from autoimmune diabetes. (a) Kinetics of development of glycosuria in female NOD mice that were untreated (UT) or fed (twice, at 9th and 10th week of age) titrated doses of CP (as indicated) or vehicle control (average of three independent experiments with five mice per group in each experiment, n=15 per experimental group). Urine glucose was measured twice weekly using Diastix, as described in Methods. (b) Average % incidence (±s.d.) of diabetes in untreated, vehicle (veh)-fed, and CP (10 μg)-fed NOD mice, from three independent experiments (n=15). (c) Graphical analysis of dose dependence of CP (from panel a) in mediating protection from diabetes in NOD mice after 20 weeks, based on three independent experiments with five mice per group in each experiment (n=15). (d) Pancreatic histology of NOD untreated and vehicle or CP fed (as indicated) at 20 weeks of age. Pancreata were stained with hematoxylin and eosin and analyzed at original magnification × 200 (upper panel) and original magnification × 400 (lower panel). Histology data are representative of 14 different islets from four CP 10 μg fed mice. **P=0.01.
Effect of CP on T cells is not a result of Treg cell function
Treg cells have been defined as a subset of potent immune response regulators that express CD4, CD25, and Foxp3. Both quantitative and qualitative changes in Tregs have been implicated as an important event in the development of T1D in NOD mice. We considered that one possible explanation for our observations is enhancement of Tregs, either as a numerical increase, or alternatively, as an enhancement of their suppressive function. However, we found in our fluorescence-activated cell sorting (FACS) analysis of CD4+Foxp3+ frequencies and total numbers in the PLN, pancreas, and spleen that there was no discernable difference in frequency or total numbers after 2 weeks of feeding (Supplementary Figure S3 online). We also utilized a common assay to test ex vivo, the suppressive function of Tregs, purified from the PLN of mice that were treated in vivo by feeding vehicle or doses of CP, and found no differences in their suppressive function (Supplementary Figure S3 online).
Multiple subsets of Treg cells have been identified, including cells with regulatory function that do not express Foxp3. In order to rule out the possibility that any T-cell population was conferring the suppression of T-cell proliferation, we utilized an in vitro proliferation assay. Naive NOD mice were fed with CP or vehicle, and PLN of the fed mice were harvested after 2 days. PLN cells were irradiated and cocultured in vitro with naive CD4+CD25− T cells (or T effectors) purified from untreated NOD (polyclonal T effectors; Figure 3a, left panel) or BDC2.5 (antigen-specific T effectors; Figure 3a, right panel) in the presence of soluble αCD3 antibody. T-cell proliferation was assessed by [3H]-thymidine incorporation. In accordance with our findings in vivo, proliferation of T cells was also inhibited by PLN cells in vitro (Figure 3a). Similar inhibition of BDC2.5 T effectors was observed when stimulated with BDC2.5 mimetope peptide (1040–31; 5 μg ml−1) (Supplementary Figure S4 online). However, the inhibition was to a modest degree as compared with our in vivo data. We reasoned this could be due to stronger external signals to T cells from αCD3 or exogenous peptide stimulation, or could be due to in vivo neuronal effects absent in an in vitro system.

Attenuation of T-cell proliferation is dependent on CD11b+F4/80+ macrophages. (a) Prediabetic non-obese diabetic (NOD) mice (n=3) were with fed Capsaicin (CP; 10 μg) or vehicle (veh) control, and 2 days later pancreatic lymph nodes (PLNs) were harvested and cocultered (5 × 104) with CD4+CD25− (1 × 105) cells purified from naive untreated NOD (left panel) or naive untreated NOD.BDC2.5 (right panel) at the indicated ratios in the presence of soluble αCD3 antibody (0.5 μg ml−1) for 3 days. Graph indicates average c.p.m. (±s.d.), as measured by [3H]-thymidine incorporation for the last ∼20 h of culture. Data are representative of at least three independent experiments. (b) Same experimental design as in panel a comparing whole PLN to T-depleted PLN, CD11b-depleted PLN, or CD11b-enriched PLN, respectively. T cells were depleted using CD90.2 microbeads and antigen-presenting cells were depleted using CD11b microbeads (Miltenyi). Data are representative of two independent experiments with three mice per group. (c) NOD or CD11b.DTR/NOD was injected with diphtheria toxin (DT) to ablate macrophages or phosphate-buffered saline (PBS) control as described in Methods. DT- or PBS-treated PLNs were used in proliferation assay as in panel a. Fluorescence-activated cell sorting (FACS) plots (left panel) indicate the CD11b and CD11c expression in CD11b.DTR/NOD PLN treated with PBS or DT 48 h prior, as indicated. Numbers on FACS plots indicate the percent in each quadrant. Panel d indicates the average c.p.m. (±s.d.) of [3H]-thymidine incorporation in the treated groups, as indicated (three mice per group). Data are representative of two independent experiments. *P=0.05, **P=0.01, ***P=0.001. Te, treated; NS, nonsignificant.
To further test the role of Treg cells, we depleted PLN cells of all T cells (T-depleted PLN) using microbeads and tested their ability to induce proliferation of naive T effectors in vitro. Depletion of T cells did not abrogate the suppressive ability of the PLN cells in CP 10 μg fed mice when compared with vehicle (Figure 3b). These data clearly indicates that suppressor T cells are not responsible for the abrogation of proliferation as T-depleted PLN cells still remain suppressive. In light of these results we turned our attention to another potent immunological subset, APC, which are known to express VR1.
Attenuation of T cells by CP is dependent on the APC compartment
APCs such as DCs, macrophages, and B cells are a key cell type of the immune system, linking innate, and adaptive immunity. They can either provoke or alleviate an autoimmune disease.12, 23, 24, 25, 26 Phenotypic analysis of APC populations present in the PLN revealed several distinct populations. We used the markers CD11b and CD11c to identify three APC subsets that expressed high levels of these markers, CD11bhi, CD11b+CD11c+, and CD11c+ (Supplementary Figure S5 online). FACS analysis with a panel of antibodies allowed for further characterization of these populations. The CD11bhi population expressed a low level of CD11c, an intermediate level of MHCII, and all highly expressed the macrophage marker F4/80 indicating that these cells are macrophages. The CD11bhiF4/80+ macrophages did not express Gr-1 or CD40, but did express the costimulatory molecules CD80 and CD86 and the inhibitory molecule PD-L1. The CD11b+CD11c+-double positive population expressed a high level of CD11c and an intermediate level of CD11b. These cells also expressed a high level of MHCII in addition to CD40, CD80, CD86, and PD-L1, but were negative for Gr-1. We define these cells as DCs. The final population we analyzed was a CD11c+CD11b− population. This population appeared to be heterogeneous in nature, as indicated by two distinct levels of MHCII. There was no expression of F4/80 on these cells, but analysis of all the other markers assayed revealed two distinct populations as the CD11c+CD11b− had both positive and negative cells. The CD11c+CD11b− population was likely comprised of heterogeneous DCs of different maturation states, including immature DCs (Supplementary Figure S5 online).
To assess their role in CP-mediated suppression, we depleted the CD11b+ cells, an integrin expressed by APCs, both in vitro and in vivo. CD11b microbeads were used to deplete the CD11b+ cells in vitro, from PLN of NOD mice that had been previously fed with CP 10 μg or vehicle (Figure 3b). Interestingly, the suppressive effect of PLN cells from CP-fed mice was completely abolished after depletion of CD11b+ cells. Further, when CD11b-enriched PLN cells from vehicle and CP-fed mice were cocultured with T-effector cells, a massive inhibition of proliferation was observed (Figure 3b), thus demonstrating that CD11b+ cells are responsible for CP-mediated T-cell suppression.
As in vitro depletion of CD11b+ cells depleted the CD11b+CD11c+ DCs along with CD11b+/F480+ macrophages, we took an additional approach. We depleted the CD11bhiF4/80+ cells in vivo using the CD11b.DTR/NOD mice. These mice when treated with 25 ng of diphtheria toxin preferentially eliminate CD11bhiF4/80+ macrophages, but not the CD11b+CD11c+ DCs (Figure 3c). Pretreating the CD11b.DTR/NOD mice with diphtheria toxin 24 h prior to feeding completely abolished the suppressive effect of PLN cells from CP-fed mice (Figure 3d). These data confirmed that CD11bhiF4/80+ macrophages are the suppressor cells in PLN of CP-fed mice.
Moderate but preferential increase in CD11bhiF4/80+ population after CP feeding
The APC population conferring the protective effect of CP was clearly CD11bhiF4/80+ and so we examined the CD11bhiF4/80+ and CD11b+CD11c+ populations further. We analyzed the frequencies of the CD11bhiF4/80+ macrophages and CD11b+CD11c+ DCs in NOD mice fed with CP (10 or 25 μg) or vehicle after 48 h. Comparison of the two APC frequencies in the PLN revealed that there was a two- to three-fold increase in the frequency of CD11bhiF4/80+ population, but not in the CD11b+CD11c+, after feeding with CP 10 μg when compared with vehicle or CP 25 μg dose (Figure 4a and b). The composition of other leukocytes such as B cells and natural killer cells have been tested on days 2–7 in the PLN of NOD mice. We did not find any difference in B cells, natural killer cells, and CD11b+/CD11c+ DC population, whereas a modest but significant increase in CD11bhi cells was observed starting 2 days post-feeding in the PLN. This increase is maintained for at least 7 days, although it tapers off over time. This observed increase in macrophages is a general phenomenon and not restricted to the NOD strain, but was evident in other strains of mice such as BALB/c (Figure 4c), C57BL/6 mice shown later (Figure 5d), and RIP-OVA mice (Supplementary Figure S7 online) fed with different doses of CP or vehicle. In the RIP-OVA mice, we have noted a significant increase in CD11bhi cells in the PLN 4 days after feeding (3 days after OT-I transfer, as in Figure 1b). There was no difference in the frequency of CD11bhi cells in the RLN. This finding was also reflected in the total numbers of CD11bhiF4/80+ macrophages. Interestingly, the dose of CP at which the CD11bhiF4/80+ cells peaked differed between strains; 5 μg of CP in BALB/c mice and 10 μg of CP in NOD and C57BL/6. The reason for this presently remains unclear, but may be a consequence of differences in VR1 expression or affinity between the strains or other unspecified strain differences.

CD11bhi macrophages in pancreatic lymph nodes (PLNs) attain regulatory phenotype after oral administration of Capsaicin (CP). (a) Representative fluorescence-activated cell sorting (FACS) plots of naive non-obese diabetic (NOD) fed vehicle (veh) or titrated doses of CP (as indicated), and 2 days later FACS analysis of PLN was performed for CD11b and CD11c expression. Numbers in quadrants indicate percent cells in each (n=3). Data are representative of three independent experiments. (b) Identical experiment as in panel a comparing CD11bhi frequency in naive NOD or (c) BALB/c after feeding vehicle or titrated doses of CP, as indicated (n=4–5). Graph indicated average % (±s.d.) CD11bhi in PLN. Data are representative of many independent experiments. (d) Prediabetic recipient NOD mice (n=4–5) were fed CP (10 μg) or vehicle control, and 2 days later FACS analysis of PLN was performed on CD11bhiF4/80+ macrophages and CD11b+CD11c+ dendritic cell (DC) for PD-L1 expression. Representative of three independent experiments. (e) Prediabetic NOD mice were fed CP (at doses indicated) or vehicle at 9 and 10 weeks of age. At 11 weeks of age, PLN were harvested and intracellular cytokine staining was performed ex vivo on CD11bhiF4/80+ macrophages and CD11b+CD11c+ DC for interleukin (IL)-10. Representative of three independent experiments. Graph indicates % (±s.d.) IL-10+ of CD11bhi MØ or CD11b+CD11c+ DC population as indicated. (f) Prediabetic NOD mice (n=3) were fed CP (10 μg) or vehicle control, and 2 days later PLN were harvested and cocultured (5 × 104) with CD4+CD25− (1 × 105) cells purified from naive NOD.BDC2.5 in the presence of soluble aCD3 (0.5 μg ml-1) +/− αIL-10 and αIL-10R (1 μg ml−1 each) or immunoglobin-G1 (IgG1) and IgG2b (1 μg ml−1 each). Graph indicates average c.p.m. (±s.d.), as measured by [3H]-thymidine incorporation for the last ∼20 h of culture. Data are representative of two independent experiments. *P=0.05, **P=0.01. NS, nonignificant; PBS, phosphate-buffered saline.

Suppression of T-cell proliferation is dependent on expression of VR1. (a and b) rat insulin promoter (RIP)-ovalbumin (OVA) and RIP-OVA/VR1−/− (n=3–4 per group) were fed Capsaicin (CP; 10 μg or 25 μg) or vehicle (veh), and 24 h later received adoptive transfer of CFSE-labeled OT-I CD8+ cells intravenous Four days post-transfer pancreatic lymph nodes (PLN) were harvested and analyzed by fluorescence-activated cell sorting (FACS) for CFSE dilution. Representative histograms of CFSE dilution (right panel) and average % (±s.d.) of divided OT-I T cells out of total the OT-I transferred population (left panel) of RIP-OVA recipients (a) and RIP-OVA/VR1−/− (b). Representative of at least two independent experiments. (c) RIP-OVA/VR1−/− mice (n=4–5 per group) were lethally irradiated and 24 h later received CD45.1 C57BL/6 bone marrow (BM) as described in Methods. After reconstitution (6 weeks), RIP-OVA/VR1−/− BM chimeras were treated as in panel a. Graph indicates average % (±s.d.) of divided OT-I cells out of the total OT-I transferred population. Representative of two independent experiments. (d) C57BL/6 and VR1−/− were fed vehicle or titrated doses of CP (as indicated), and 2 days later PLN were harvested and FACS analysis was performed for CD11b and CD11c expression. Graph indicated average % (±s.d.) CD11bhi in PLN. Data are representative of two independent experiments. (e) C57BL/6 and VR1−/− mice were fed CP (10 μg) or vehicle (n=3 per group), and 2 days later PLN were harvested, separated into single-cell suspension, irradiated, and cocultured in vitro at the indicated ratio with naive CD4+CD25− cells purified from OT-II T-cell antigen receptor transgenic mice and OVA peptide 323–339. Proliferation was measured as previously described. Representative of two independent experiments. *, P=0.05, ***, P=0.001. NS, nonsignificant; VR1, vanilloid receptor 1.
CD11bhiF4/80+ macrophages from CP-fed mice have regulatory phenotype
The marked inhibition by CD11b-enriched PLN cells from CP-fed mice (Figure 3b) suggested qualitative differences in macrophage function in addition to their increase in frequency (Figure 4a–c). Engagement of VR1 on DCs is known to modulate the expression of antigen-presenting and costimulatory molecules.4 Thus, we determined whether the same holds true when CP is administered orally. We compared surface expression of MHCII, CD86, and PD-L1 on the APC populations in PLN of fed mice by flow cytometry. PD-L1, a coinhibitory molecule, was upregulated specifically in CD11bhiF4/80+ macrophages from CP-fed mice, but not MHCII or CD86 (Figure 4d and Supplementary Figure S6a, b online). However, no difference was observed in the expression levels of these markers on the CD11b+CD11c+ population. These data suggest that oral administration of CP alters the activation status of the macrophages.
In addition to providing inhibitory signals via their cell surface markers, APCs can also secrete a myriad of cytokines that influence immunity. The levels of cytokines produced by these small populations of PLN cells were below the limit of detection by both ELISA and multiplex assays. Thus, we performed intracellular cytokine staining of the CD11bhiF4/80+ and CD11b+CD11c+ populations. In particular, we were interested in interleukin (IL)-10, an immune-suppressive cytokine with broad anti-inflammatory properties essential for maintaining immune homeostasis in the gut-associated lymphoid tissue.27 We observed that 4 days after CP administration there was a two-fold increase in IL-10 production in CD11bhiF4/80+ macrophages ex vivo without in vitro stimulation (Figure 4e and Supplementary Figure S6c online). In contrast, the percentage of cells producing IL-10 in the CD11b+CD11c+ remained unaltered after feeding with vehicle, 10 μg or 25 μg of CP (Figure 4e). Therefore, feeding with 10 μg of CP enhances a multifactorial regulatory phenotype in CD11bhiF4/80+ macrophages by upregulation of PD-L1 and IL-10 expression.
CP-mediated regulation of T cells is IL-10 dependent
The present studies have described an increase in IL-10-producing macrophages in the PLN, and these macrophages are required for regulation of T-cell responses. Having established the essential role of macrophages, we next asked if their IL-10 production directly contributes to T-cell suppression. PLN of NOD mice fed with CP 10 μg or vehicle were cocultured with naive T effectors in the presence of antibodies neutralizing IL-10 and IL-10 receptor in our in vitro T-cell proliferation assay. Neutralizing IL-10 completely reversed the suppressive effect of PLN cells from CP 10 μg fed mice compared with the isotype and vehicle controls (Figure 4f). This finding indicates that CP-mediated immunoregulation of T-cell proliferation is IL-10 dependent.
Inhibition of T-cell proliferation is dependent on the expression of VR1
To determine whether VR1 is required for CP-mediated suppression of autoreactive T cells, we crossed RIP-OVA and VR1−/− mice to generate RIP-OVA/VR1−/− mice, a model of T1D that is devoid of functional VR1 but expresses OVA in the pancreas. We fed RIP-OVA and RIP-OVA/VR1−/− mice with CP (10, 25 μg) and vehicle, and assayed for OT-1 proliferation in vivo. In these mice, CP-mediated inhibition of OT-1 proliferation was completely abrogated at any dose tested, indicating that the inhibition of autoreactive T-cell proliferation is dependent on the expression of VR1 (Figure 5a and b).
We next questioned if enhancement in frequency of macrophage is VR1 dependent. In the absence of VR1−/− mice, the macrophage frequency in PLN cells failed to increase at any dose of CP tested, suggesting that engagement of VR1 enhances the frequency of macrophages in PLN (Figure 5d).
As frequency of CD11bhiF4/80+ macrophages in PLN remained unchanged after feeding with CP in VR1−/− mice, we next tested if a regulatory effect of PLN cells is also abolished in the absence of VR1. For this, we set up an ex-vivo antigen–specific T-cell proliferation assay. OT-II CD4+ T cells were cocultured with PLN cells from RIP-OVA or RIP-OVA/VR1−/− mice fed with CP. After 3 days of culture, OT-II proliferation was assessed by [3H]-thymidine incorporation. Proliferation of OT-II cells was significantly inhibited when stimulated with PLN cells from RIP-OVA mice. In contrast, no inhibition was evident when stimulated with PLN from RIP-OVA/VR1−/− mice (Figure 5e). Together, these data indicate that CP-mediated suppression of autoreactive T-cell proliferation requires VR1 expression.
VR1 expression is required on APCs
VR1 is expressed on sensory neurons and APCs.4, 28 VR1 expressing sensory neurons have been reported to innervate the gut.29, 30, 31, 32 VR1 stimulation is known to secrete neuropeptides like substance P (SP) or calcitonin gene-related peptide from sensory neurons, which can act as immune modulators. On the other hand, we have reported that DCs and macrophages express VR1, and engagement of VR1 on DCs has powerful immune consequences.4, 7 We have also tested by semiquantitative PCR that CD11bhi macrophages isolated from PLN and mesenteric lymph node express VR1 transcript (data not shown). Thus potentially, orally delivered CP can interact with VR1 on sensory neurons lining the gut, which in turn can influence the immune system. Alternatively, CP could be sensed directly by APCs in the gastrointestinal tract. Having established that VR1 function is required for CP-mediated T-cell suppression, we next examined whether this immuno-modulatory effect is due to neuronal cells or immune cells or both. In order to tease out the mechanism we generated bone marrow (BM) chimeras. Lethally irradiated RIP-OVA/VR1−/− mice were reconstituted with wild-type CD45.1 BM from C57BL/6 to generate chimeric mice that were VR1 sufficient in myeloid-derived cells and VR1 deficient in sensory neurons. After 4 weeks post-BM transfer, the recipient mice had reached at least 90% CD45.1 chimerism. The DRG neurons from these mice were found to be CP insensitive, confirming that immune cells were VR1+/+ but neuronal cells were VR1−/−. We then tested the proliferation of OT-I cells in these mice after feeding. If inhibition of T-cell proliferation is observed to a significant extent in the VR1+/+ → VR1−/− BM chimeras, we can conclude that CP is being sensed directly by the immune system through the VR1+/+ myeloid compartment. As illustrated in Figure 5c, the proliferation of OT-I cells in VR1+/+ → VR1−/− chimeric mice was inhibited to a significant extent in the CP 10 μg fed group in comparison with vehicle. The inhibition was substantial but not as strong as observed in RIP-OVA mice (Figures 5a and 1b). We also did not observe inhibition in the CP 25 μg group as seen in RIP-OVA mice. This indicates that VR1-sufficient immune cells at 10 μg dose can suppress autoreactive T-cell activation independent of sensory neurons. Engagement of VR1 on sensory neurons likely strengthens the suppressive effect, as inhibition of T-cell proliferation is not fully restored by VR1+/+ → VR1−/− BM chimerism.
Discussion
Our observations here show how a commonly used nutritional factor, CP, can modulate gut-mediated immune tolerance and autoimmunity: (i) orally administered CP induces subtle changes in the immune parameters in PLN that leads to suppression of an ongoing immune response and prevention from autoimmune diabetes; (ii) this is a general phenomenon and not restricted to any particular system or strain of mice; (iii) the protective dose is restricted to 10 μg and higher doses, i.e., 25 or 50 μg, does not lead to any immune suppression; (iv) suppression is specifically mediated through CD11bhiF4/80+ cells in the PLN; (v) suppression is IL-10 dependent; and (vi) the suppressive effect is derived from at least two distinct sources, one is the myeloid compartment and the other is a non-myeloid compartment presumably the neuronal components.
We emphasize based on our observations that the suppressive effect of CP is dependent on oral route, which specifically impinge on the gut lymph nodes. CP when administered by other routes, e.g., intra-dermal or subcutaneous did not show any immunosuppressive effect (data not shown). We were unable to administer by intravenous route, as LD50 of CP in mice is 10–12 μg for intravenous injection.
The dose–response of CP is surprisingly narrow. We note repeatedly that this phenomenon is tightly dose-restricted. Lower doses of CP are probably below the threshold at which differences in immunological outcome can be measured. Higher doses of CP might be toxic and perturb the gastrointestinal milieu in an inflammatory manner. It is possible that at higher dose, CP can also be sensed by sensory neurons or even T cells, which may counteract the suppressive phenotype observed at the protective dose.
Our data clearly show that VR1 engagement is critical for enhancement of CD11bhi/F480+ macrophage frequency and their regulatory phenotype in the gut-draining PLN after feeding with CP. These macrophages inhibit autoreactive CD4+ and CD8+ T-cell proliferation and function. The suppressive effect of CP is only manifested in gut macrophages, specifically in PLN, but not in other peripheral lymph nodes. Although we have observed that CD11bhi macrophages isolated from PLN and mesenteric lymph node express VR1 transcript, the possibility that expression of VR1 is different in different macrophage populations need to be examined. Conversely, the possibility that gut environment might influence suppressive effects deserves experimental enquiry. On the basis of the recent reports12, 33, 34, 35 we believe that gut-resident macrophages are profoundly anergic and fail to produce inflammatory cytokines upon stimulation and instead make IL-10. Our data are consistent with these reports. These macrophages are unique in their phenotype as compared with other lymph node macrophages and may contribute to the maintenance of intestinal tolerance in vivo. This uniqueness might be the effect of the gut microenvironment analogous to the effect of adipocytes on adipose tissue macrophages.36, 37, 38
In contrast to previous findings39 on autoimmune diabetes, our data show that administration of CP did not impact the number or the function of Foxp3 expressing Treg cells in the draining PLN or in the pancreatic tissue, but could elicit a number of suppressor factors such as IL-10 and PD-L1 in a discreet population of CD11bhiF4/80+ macrophages found in the same lymph node. The enhancement of macrophage frequency and difference in their phenotypic markers (Figure 4) were undoubtedly modest, but sufficient to significantly suppress an ongoing immune response. Interestingly, depletion of these macrophages in vivo in CD11b.DTR/NOD mice (Figure 3d) and in vitro by neutralizing secreted IL-10 by antibodies to IL-10 and IL-10 receptor restored T-cell proliferation specifically in CP-treated mice. Thus our data indicate that phenotypic changes in macrophage population, specifically IL-10, are critical for T-cell tolerance in PLN. Further questions remain regarding the mechanism. Even though we did not detect any Foxp3 Tregs in vivo, we cannot exclude the possibility that FoxP3-negative Treg cells can be induced by these macrophages. We have observed that after 14 days of CP feeding, CD4+ T cells in the spleen and PLN exhibit Tr-1-like suppressor T-cell phenotype, expressing IL-10, and can transfer protection from T1D in untreated NOD mice (E. Nevius and S. Basu, unpublished observation). Mechanism of Tr-1 generation will help to elucidate how the subtle changes in macrophage population can lead to such marked inhibition of effector T-cell function and protection from T1D.
Histological examination of the islets from mice fed with CP reveal that most of the islets have no insulitis. But even in the infiltrated islets, the accumulation of inflammatory cells is restricted to the periphery. It is likely that the suppressor macrophages contribute to resolution of insulitis. Recent studies suggest that innate immune cells and inflammatory mediators largely contribute to the induction, amplification, stabilization, and maintenance of insulitis. Complete resolution of insulitis is observed in many individuals who had initially developed mild insulitis, but do not show any histological signs of insulitis at a later stage. Resolution of inflammation is an active process involving proresolution mediators found in many tissues.40, 41, 42, 43 Our observations altogether suggest that CP feeding at 9 weeks of age either inhibits maintenance of insulitis or actively participate in resolution of insulitis. CP might possess dual anti-inflammatory and proresolution activity that needs further investigation.
VR1 and pancreas have an intimate relation. VR1-expressing neurons innervate the pancreas.44 According to Razavi et al,45 ablation of VR1-expressing neurons in the pancreas can protect NOD mice from T1D. These authors have speculated that a fine balance between the neuropeptides and β-cell stress in the pancreas determines disease outcome. In their system, CP treatment in neonates ablates entire sensory nerves and probably modulates the immune cells as well. Our data indicate the contrary. In VR1−/− mice we observed unimpeded T-cell activation after oral administration of CP, which clearly underlines the significance of VR1 engagement for T-cell tolerance and autoimmunity. We demonstrate that engagement of VR1 on the immune cells is necessary and sufficient for attenuating effector T-cell function, but at the same time appreciate the fact that neuro-immune crosstalk via VR1 might strengthen this response.
Finally, CP is a widely used nutritional factor. A recent report by Kang et al,38 suggest that dietary CP can attenuate metabolic dysregulation by suppressing adipose tissue inflammatory responses. Even though our present study does not focus on the nutritional aspects of CP, it clearly demonstrates the beneficial effects of CP on T-cell tolerance and autoimmunity. Our study substantiates the notion that nutritional factors can have profound effects on immune response and leans to the idea that endogenous ligands of VR1 may be contributing to immune regulation in the gut. In conclusion, our study identifies a fundamental relation between nutrients and sensory neurons directly regulating gut-mediated immune response.
Methods
Mouse strains. Six- to nine-week-old NOD/Lt, BALB/c, C57BL/6, TRVP1−/−, RIP-OVAhi NOD/CD11b DTR, and NOD.BDC2.5 TCR Tg mice were obtained from the Jackson Laboratory (Bar Harbor, ME). Six- to -8-week-old OT-II TCR Tg RAG−/− mice were obtained from Taconic (Germantown, NY). CD45.1+OT-I/RAG−/− expressing the CD45.1 congenic marker was a gift from Leo Lefrancois (University of Connecticut Health Center) and bred at the University of Connecticut Health Center (UCHC). All animals were housed in the Center for Laboratory Animal Care at UCHC. RIP-OVAhi and TRPV1−/− strains were backcrossed for 8–10 generations to generate RIP-OVA/VR1+/+ strain and housed at UCHC. All experiments were conducted under the guidelines approved by UCHC Animal Care Committee.
Antibodies and flow cytometry. Fluorescently labeled antibodies were purchased from Biolegend (San Diego, CA), BD (San Jose, CA), or eBioscience (San Diego, CA). FACS samples were analyzed on BD LSRII or BD FACSCalibur instruments (Becton Dickinson, San Jose, CA), and data were examined using Flowjo Version 8.8.6 (Tree Star, Ashland, OR).
Procedure for oral administration of CP and diabetes assessment. Mice were anesthetized with ketamine (2.0 mg kg−1) and xylazine (0.1 mg kg−1). Vehicle control or CP doses were gavaged 100 μl volume using an 18-gauge feeding needle (Popper & Sons, New Hyde Park, NY). Diabetes was measured by monitoring urine glucose using Diastix reagent stips (Bayer, Elkhart, IN). Mice were considered diabetic after two consecutive readings of >2,000 mg dl−1 of urine glucose.
In vitro T-cell assays and culture. Primary lymphocyte cultures were maintained in RPMI 1640 medium supplemented with 5% fetal bovine serum, 2 mM L-gluatamine, 1 mM non-essential amino acids, 1 mM sodium pyruvate, 50 units ml−1 penicillin/streptomycin, and 50 μM β-mercaptoethanol, all purchased from Gibco/Invitrogen (Carlsbad, CA). For in vitro T-cell proliferation assays, prediabetic NOD mice were fed CP (10 μg) or vehicle control, and 2 days later PLN were harvested and cocultured with CD4+CD25− cells purified from naive untreated NOD or NOD.BDC2.5 at the indicated ratios in the presence of soluble αCD3 Ab (0.5–1 μg ml−1). T cells were depleted using CD90.2 microbeads and APCs were depleted using CD11b microbeads (Miltenyi Biotech, Auburn, CA). Proliferation was measured over the last ∼20 h of a 3-day culture by [3H]-thymidine incorporation (2 μCi per well) and is represented as c.p.m. In some cases, proliferation was measured in cultures of PLN from RIP-OVA or RIP-OVA/VR1−/− mice with OT-II/Rag−/− T effectors.
Adoptive transfer experiments. CD8+ OT-I or CD4+ BDC2.5 were isolated from CD45+ OT-I TCR Tg RAG−/− or NOD.BDC2.5 TCR Tg mice, respectively. Spleens and LN were harvested from donor mice and enriched for CD8+ or CD4+ cells using anti-CD8 or anti-CD4 microbeads (Miltenyi Biotech) according to the manufacturer's instructions. Adoptive transfer recipients (RIP-OVA or NOD) were fed vehicle or titrated doses of CP, and 24 h later received between 0.5 × 106 and 1.0 × 106 CFSE (Molecular Probes)-labeled TCR Tg T cells from the appropriate strain of mice (OT-I or BDC2.5) via retro-orbital injection. Three to four days post-adoptive transfer, PLN were harvested and analyzed by FACS for dilution of CFSE and expression of activation markers. The adoptively transferred T cells were distinguished by staining with antibodies to CD8, and congenic marker CD45.1 or in some cases with CD8 and Kb pentamer associated with the OVA-derived peptide SIINFEKL (see Supplementary Figure S2 online). In RIP-OVA mice, RLNs were also harvested to analyze CFSE dilution.
Intracellular cytokine staining. For T-cell intracellular cytokine staining, PLNs were harvested from NOD mice 4 days after feeding, separated into single-cell suspension, and cultured in complete RPMI medium and PMA (200 ng ml−1) plus ionomycin (1 μg ml−1; for T cells) in the presence of brefeldin A for 4 h and incubated at 37 μC. After incubation, cells were fixed with 1.5% formaldehyde and permeablized in 100% methanol. Staining was performed on fixed/permeabilized cells that were stained using fluorescent antibodies. For APC intracellular cytokine staining, PLNs were harvested from NOD mice 48 h after feeding, mechanically separated into single-cell suspension and immediately fixed, permeabilized, and stained as described above.
Bone marrow chimeras. Six- to 8-week-old RIP-OVA/VR1−/− mice received ∼1,000 rads in a single dose of γ-irradiation. Twenty-four hours later via retro-orbital injection, irradiated hosts are adoptively transferred with 2 × 106 BM cells harvested from femurs and tibias of CD45.1+C57BL/6 wild-type mice. After 4 weeks, BM chimeric mice were bled via tail bleed to check for BM reconstitution by FACS.
Histopathology. Pancreata were fixed in formalin, embedded in paraffin, sectioned, and stained with hematoxylin/eosin.
Statistical analyses. Statistical comparisons were calculated by Student's t-test (unpaired, two-tailed) using GraphPad Prism Version 5.0 (GraphPad Software, La Jolla, CA). Significance was set at P<0.05.
References
Tominaga, M. et al. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 21, 531–543 (1998).
Cesare, P., Moriondo, A., Vellani, V. & McNaughton, P.A. Ion channels gated by heat. Proc. Natl. Acad. Sci. USA 96, 7658–7663 (1999).
Caterina, M.J. et al. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 288, 306–313 (2000).
Basu, S. & Srivastava, P. Immunological role of neuronal receptor vanilloid receptor 1 expressed on dendritic cells. Proc. Natl. Acad. Sci. USA 102, 5120–5125 (2005).
Bodo, E. et al. A hot new twist to hair biology: involvement of vanilloid receptor-1 (VR1/TRPV1) signaling in human hair growth control. Am. J. Pathol. 166, 985–998 (2005).
Lu, T., Newton, C., Perkins, I., Friedman, H. & Klein, T.W. Role of cannabinoid receptors in Delta-9-tetrahydrocannabinol suppression of IL-12p40 in mouse bone marrow-derived dendritic cells infected with Legionella pneumophila. Eur. J. Pharmacol. 532, 170–177 (2006).
Beltran, J., Ghosh, A.K. & Basu, S. Immunotherapy of tumors with neuroimmune ligand capsaicin. J. Immunol. 178, 3260–3264 (2007).
Flemming, A. Red chillies tackle tumours. Nat. Rev. Drug Discov. 6, 269 (2007).
Steinman, R.M., Hawiger, D. & Nussenzweig, M.C. Tolerogenic dendritic cells. Annu. Rev. Immunol. 21, 685–711 (2003).
Uematsu, S. et al. Regulation of humoral and cellular gut immunity by lamina propria dendritic cells expressing Toll-like receptor 5. Nat. Immunol. 9, 769–776 (2008).
Iwasaki, A. Mucosal dendritic cells. Annu. Rev. Immunol. 25, 381–418 (2007).
Denning, T.L., Wang, Y.C., Patel, S.R., Williams, I.R. & Pulendran, B. Lamina propria macrophages and dendritic cells differentially induce regulatory and interleukin 17-producing T cell responses. Nat. Immunol. 8, 1086–1094 (2007).
Sun, C.M. et al. Small intestine lamina propria dendritic cells promote de novo generation of Foxp3 T reg cells via retinoic acid. J. Exp. Med. 204, 1775–1785 (2007).
Mowat, A.M. Anatomical basis of tolerance and immunity to intestinal antigens. Nat. Rev. Immunol. 3, 331–341 (2003).
Kelsall, B.L. & Strober, W. Distinct populations of dendritic cells are present in the subepithelial dome and T cell regions of the murine Peyer′s patch. J. Exp. Med. 183, 237–247 (1996).
Turley, S.J., Lee, J.W., Dutton-Swain, N., Mathis, D. & Benoist, C. Endocrine self and gut non-self intersect in the pancreatic lymph nodes. Proc. Natl. Acad. Sci. USA 102, 17729–17733 (2005).
Horwitz, M.S. et al. Diabetes induced by Coxsackie virus: initiation by bystander damage and not molecular mimicry. Nat. Med. 4, 781–785 (1998).
Schmid, S. et al. Delayed exposure to wheat and barley proteins reduces diabetes incidence in non-obese diabetic mice. Clin. Immunol. 111, 108–118 (2004).
Wen, L. et al. Innate immunity and intestinal microbiota in the development of Type 1 diabetes. Nature 455, 1109–1113 (2008).
Hartmann, B., Bellmann, K., Ghiea, I., Kleemann, R. & Kolb, H. Oral insulin for diabetes prevention in NOD mice: potentiation by enhancing Th2 cytokine expression in the gut through bacterial adjuvant. Diabetologia 40, 902–909 (1997).
Chatenoud, L., Thervet, E., Primo, J. & Bach, J.F. Anti-CD3 antibody induces long-term remission of overt autoimmunity in nonobese diabetic mice. Proc. Natl. Acad. Sci USA 91, 123–127 (1994).
Kurts, C. et al. Constitutive class I-restricted exogenous presentation of self antigens in vivo. J. Exp. Med. 184, 923–930 (1996).
Clare-Salzler, M.J., Brooks, J., Chai, A., Van Herle, K. & Anderson, C. Prevention of diabetes in nonobese diabetic mice by dendritic cell transfer. J. Clin. Invest. 90, 741–748 (1992).
Feili-Hariri, M. et al. Immunotherapy of NOD mice with bone marrow-derived dendritic cells. Diabetes 48, 2300–2308 (1999).
Zhou, Y. et al. Oral tolerance to food-induced systemic anaphylaxis mediated by the C-type lectin SIGNR1. Nat. Med. 16, 1128–1133 (2010).
Manicassamy, S. et al. Activation of beta-catenin in dendritic cells regulates immunity versus tolerance in the intestine. Science 329, 849–853 (2010).
Moore, K.W., de Waal Malefyt, R., Coffman, R.L. & O′Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 19, 683–765 (2001).
Caterina, M.J. et al. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 389, 816–824 (1997).
Patterson, L.M., Zheng, H., Ward, S.M. & Berthoud, H.R. Vanilloid receptor (VR1) expression in vagal afferent neurons innervating the gastrointestinal tract. Cell Tissue Res. 311, 277–287 (2003).
Robinson, D.R., McNaughton, P.A., Evans, M.L. & Hicks, G.A. Characterization of the primary spinal afferent innervation of the mouse colon using retrograde labelling. Neurogastroenterol Motil. 16, 113–124 (2004).
Ward, S.M., Bayguinov, J., Won, K.J., Grundy, D. & Berthoud, H.R. Distribution of the vanilloid receptor (VR1) in the gastrointestinal tract. J. Comp. Neurol. 465, 121–135 (2003).
Zhang, L., Jones, S., Brody, K., Costa, M. & Brookes, S.J. Thermosensitive transient receptor potential channels in vagal afferent neurons of the mouse. Am. J. Physiol. Gastrointest. Liver Physiol. 286, G983–991 (2004).
Hirotani, T. et al. The nuclear IkappaB protein IkappaBNS selectively inhibits lipopolysaccharide-induced IL-6 production in macrophages of the colonic lamina propria. J. Immunol. 174, 3650–3657 (2005).
Kamada, N. et al. Abnormally differentiated subsets of intestinal macrophage play a key role in Th1-dominant chronic colitis through excess production of IL-12 and IL-23 in response to bacteria. J. Immunol. 175, 6900–6908 (2005).
Smythies, L.E. et al. Human intestinal macrophages display profound inflammatory anergy despite avid phagocytic and bacteriocidal activity. J. Clin. Invest. 115, 66–75 (2005).
Kang, J.H., Kim, C.S., Han, I.S., Kawada, T. & Yu, R. Capsaicin, a spicy component of hot peppers, modulates adipokine gene expression and protein release from obese-mouse adipose tissues and isolated adipocytes, and suppresses the inflammatory responses of adipose tissue macrophages. FEBS Lett. 581, 4389–4396 (2007).
Kang, J.H. et al. Dietary capsaicin reduces obesity-induced insulin resistance and hepatic steatosis in obese mice fed a high-fat diet. Obesity (Silver Spring) 18, 780–787 (2010).
Kang, J.H. et al. Dietary capsaicin attenuates metabolic dysregulation in genetically obese diabetic mice. J. Med. Food 14, 310–315 (2011).
You, S. et al. Key role of the GITR/GITRLigand pathway in the development of murine autoimmune diabetes: a potential therapeutic target. PLoS One 4, e7848 (2009).
Eizirik, D.L., Sandler, S. & Palmer, J.P. Repair of pancreatic beta-cells. A relevant phenomenon in early IDDM? Diabetes 42, 1383–1391 (1993).
Levy, B.D., Clish, C.B., Schmidt, B., Gronert, K. & Serhan, C.N. Lipid mediator class switching during acute inflammation: signals in resolution. Nat. Immunol. 2, 612–619 (2001).
Serhan, C.N., Chiang, N. & Van Dyke, T.E. Resolving inflammation: dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 8, 349–361 (2008).
Eizirik, D.L., Colli, M.L. & Ortis, F. The role of inflammation in insulitis and beta-cell loss in type 1 diabetes. Nat. Rev. Endocrinol. 5, 219–226 (2009).
Wick, E.C. et al. Transient receptor potential vanilloid 1, calcitonin gene-related peptide, and substance P mediate nociception in acute pancreatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 290, G959–969 (2006).
Razavi, R. et al. TRPV1+ sensory neurons control beta cell stress and islet inflammation in autoimmune diabetes. Cell 127, 1123–1135 (2006).
Acknowledgements
We thank Dr Qian Wu for histopathological analysis, Dr Lynn Puddington for providing us with OT-II mice, Jaqueline Beltran for technical assistance, and all other members of our laboratory for discussion and evaluation of experiments.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declared no conflict of interest.
Additional information
SUPPLEMENTARY MATERIAL is linked to the online version of the paper
Supplementary information
Rights and permissions
About this article
Cite this article
Nevius, E., Srivastava, P. & Basu, S. Oral ingestion of Capsaicin, the pungent component of chili pepper, enhances a discreet population of macrophages and confers protection from autoimmune diabetes. Mucosal Immunol 5, 76–86 (2012). https://doi.org/10.1038/mi.2011.50
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mi.2011.50
This article is cited by
-
Pancreatic draining lymph nodes (PLNs) serve as a pathogenic hub contributing to the development of type 1 diabetes
Cell & Bioscience (2023)
-
New Insights on the Role of TRP Channels in Calcium Signalling and Immunomodulation: Review of Pathways and Implications for Clinical Practice
Clinical Reviews in Allergy & Immunology (2021)
-
Modulation of M1/M2 polarization by capsaicin contributes to the survival of dopaminergic neurons in the lipopolysaccharide-lesioned substantia nigra in vivo
Experimental & Molecular Medicine (2018)
-
Role of Mast Cells and C-Sensory Fibers in Concanavalin A-Induced Paw Edema in Two Rat Strains
Inflammation (2015)
-
TRPV1 Properties in Thoracic Dorsal Root Ganglia Neurons are Modulated by Intraperitoneal Capsaicin Administration in the Late Phase of Type-1 Autoimmune Diabetes
Cellular and Molecular Neurobiology (2013)
