
Abstract
Expressed by squamous mucosal keratinocytes, calprotectin is a complex of two EF-hand calcium-binding proteins of the S100 subfamily (S100A8 and S100A9) with significant antimicrobial activity. Calprotectin-expressing cells resist invasion by Porphyromonas gingivalis, Listeria monocytogenes, and Salmonella enterica serovar Typhimurium (S. typhimurium). To understand the interactions between calprotectin and invasive bacteria, we studied the distribution of calprotectin in the cytoplasm of TR146 epithelial cells. In response to L. monocytogenes, calprotectin mobilized from a diffuse cytoplasmic distribution to a filamentous pattern and colocalized with the microtubule network. Listeria more frequently invaded cells with mobilized calprotectin. Calprotectin mobilization was listeriolysin O-dependent and required calcium (extracellular and intracellular) and an intact microtubule network. In the presence of preformed microtubules in vitro, the anti-Listeria activity of calprotectin was abrogated. To facilitate intraepithelial survival, therefore, Listeria mobilizes calprotectin to colocalize with cytoplasmic microtubules, subverting anti-Listeria activity and autonomous cellular immunity.
Similar content being viewed by others
Introduction
Calprotectin is a complex of two EF-hand calcium-binding proteins of the S100 subfamily, S100A8 and S100A9, expressed primarily in the cytoplasm of granulocytes, monocytes, and keratinocytes. 1, 2 S100 proteins are involved in cell-cycle progression, cell differentiation, and cytoskeletal–membrane interactions.3 Calcium-dependent translocation of calprotectin occurs within the cytoplasm to the cytoskeleton and plasma membrane of monocytes and epithelial cells (TR146 cells).4 In phorbol myristate acetate-activated monocytes, for example, calprotectin colocalizes with β-tubulin, which promotes release of calprotectin from the cell using a Ca2+-dependent mechanism requiring an intact microtubule network.5
Like other squamous mucosal epithelia, the gingiva expresses calprotectin in suprabasal keratinocytes.1 In oral mucosal epithelia, expression of calprotectin increases with inflammation during gingivitis, periodontitis,6 lichen planus, and diseases of viral and fungal origin such as herpes stomatitis, oral hairy leukoplakia, and candidiasis.7 In contrast, normal skin does not express calprotectin, but calprotectin is upregulated in inflammation associated with psoriasis, lupus erythematosus,8 and wound repair.9 During infection, inflammation, and wound healing, calprotectin may contribute to the innate antimicrobial defense of the keratinocyte, a form of autonomous cellular immunity.
As an antimicrobial protein complex, calprotectin shows broad spectrum activity in vitro.10, 11, 12 In epithelial cells, calprotectin expression within the cytoplasm promotes resistance to invasion by Listeria monocytogenes.13 Listeria and Salmonella enterica serovar Typhimurium (Salmonella typhimurium) are intracellular pathogens that invade epithelial cells. Listeria invasion into nonpolarized, nonphagocytic cells is mediated by surface expression of internalins A and B.14 Corresponding invasins on Salmonella are not known. After invasion, Salmonella reside and grow in endosomal vacuoles; Listeria escape the vacuole, enter the cytoplasm, and replicate.15, 16 Within the cell, bacterial routing post-invasion reflects cellular constraints, such as space and antimicrobial molecules, and species-specific virulence factors. As antimicrobial calprotectin appears to protect keratinocytes against invading bacteria in vitro, we hypothesized that successful invasion into the cytoplasm may be accompanied by altered distribution of calprotectin.
Like suprabasal squamous keratinocytes, TR146 epithelial cells normally express calprotectin in the cytoplasm.4 In response to invasion by L. monocytogenes and S. typhimurium, we analyzed calprotectin in TR146 cells. Calprotectin distribution appeared unaffected by S. typhimurium. After L. monocytogenes invasion, however, calprotectin mobilized from a diffuse to a filamentous cytoplasmic distribution, colocalizing with microtubules. Mobilization in response to L. monocytogenes required intact microtubules, extracellular and intracellular calcium, and listeriolysin O (LLO). In the presence of preformed microtubules in vitro, the anti-Listeria activity of calprotectin was inhibited.
Results
Reduced L. monocytogenes invasion into cells expressing calprotectin
To confirm that calprotectin confers autonomous cytoplasmic immunity, we generated stable calprotectin-expressing KB epithelial cell lines by transfection.13 The well-characterized intra-cellular mucosal pathogens, L. monocytogenes and S. typhimurium (S. enterica serovar Typhimurium), were tested for their ability to invade calprotectin-expressing cells in comparison to a calprotectin-negative, sham-control transfectant. KB-EGFP (sham-control) and KB-MRP8/14 (calprotectin positive) were infected with L. monocytogenes ATCC 43249, ATCC 19111, and 10403S at a multiplicity of infection (MOI) of 100 and the number of viable intracellular bacteria was enumerated using a standard antibiotic protection assay. We observed significantly fewer intracellular Listeria in calprotectin-expressing KB-MRP8/14 cells than in the sham KB-EGFP (Figure 1a). Calprotectin expression appeared to have no effect on invasion by S. typhimurium ATCC14028 or S. typhimurium SL1344 at an MOI of 100 (data not shown). The viability of intracellular Listeria within the KB transfectants was determined over time. Listeria strains were incubated with cells for 2 h, as in the standard antibiotic protection assay, followed by lysis and enumeration of viable intracellular bacteria at 3.5, 5.5, and 7.5 h. Listeria replicated within both KB-EGFP and KB-MRP8/14 cells. In calprotectin-expressing cells, Listeria strains 43249 (Figure 1b) and 19111 (Figure 1c) at MOIs of 1 and 10 consistently showed lower fold-increase in growth than in the nonexpressing transfectant. Although significantly fewer intracellular bacteria were recovered from KB-MRP8/14 than KB-EGFP at all time points (not shown), the fold-increase in growth of Listeria 43249 at an MOI 100 in calprotectin-expressing cells exceeded the nonexpressing cells (Figure 1b). For strain 19111 at an MOI of 100, growth was similar in the calprotectin-expressing and nonexpressing cells (Figure 1c). At higher intracellular densities, therefore, Listeria tended to overcome cellular restrictions and replicate more effectively in the cytoplasm.

Invasion of L. monocytogenes into KB transfectants and intracellular growth. Invasion of Listeria was determined by the antibiotic protection assay (a). Subconfluent monolayers of KB-EGFP and KB-MRP8/14 were infected for 2 h with L. monocytogenes at a MOI of 100, followed by addition of gentamicin and incubation for another 1.5 h. Monolayers were washed and lysed with distilled water to release intracellular bacteria. Bacteria were plated on tryptic-soy agar (TSA) with a spiral plater and counted after 24 h. Each experiment was performed in triplicate wells. Values are means±s.d. from a representative experiment. The experiments were repeated at least three times with similar results. The differences between KB-EGFP and KB-MRP8/14 were statistically significant (P<0.005). To determine the intracellular growth of bacteria in KB-EGFP and KB-MRP8/14, KB transfectants were incubated for 2 h with L. monocytogenes 43249 (b) or 19111 (c) at a MOI 1, 10 or 100. Gentamicin was added and cells were lysed to recover viable intracellular bacteria as described in the Methods. Shown is fold-increase in CFUs relative to 3.5 h post-inoculation. Data shown are from one representative experiment performed in triplicate and expressed as mean±s.e. Experiments with each strain and MOI were replicated as indicated: strain 43249, 2 experiments at MOI 1 and 3 experiments at MOI 10 and 100; strain 19111, 1 experiment at MOI 1 and 10, and 3 experiments at MOI 100.
Distribution of calprotectin within TR146 cells after invasion with L. monocytogenes and S. typhimurium
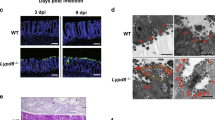
Human calprotectin (complex of S100A8 and S100A9) reacts specifically with the murine anti-human monoclonal antibody (mAb) 27E10, which does not bind monomeric or homopolymeric subunits.17 Expressed under control of native promoters, calprotectin intracellular distribution in TR146 cells was probed with mAb 27E10. In normal culture conditions, calprotectin showed diffuse cytoplasmic distribution, increasing in concentration proximal to the nucleus (Figure 2a and b). When cells were incubated with S. typhimurium at MOI of 100 for 2 h, calprotectin distribution appeared unaffected (Figure 2c and d). In contrast, cytoplasmic calprotectin mobilized into filaments in response to L. monocytogenes 43249 at MOI of 100 for 2 h (Figure 2e and f). Mobilization into filaments was detected by 1 h of incubation with L. monocytogenes 43249 and by 2 h, a maximum of 11% of cells were affected (Figure 2g). Affected cells at 2 h were undetectable at an MOI of 1 (data not shown).

Distribution of calprotectin and Listeria in the cytoplasm of TR146 epithelial cells. TR146 cells were infected with L. monocytogenes and S. typhimurium at MOI of 100 for 2 h. The cells were then fixed, permeabilized, and stained with mouse mAb 27E10-biotin (streptavidin-Alexa 568, red) against S100A8/A9 complex and DAPI (blue) for nuclear staining. Low (panels 1; scale bar is 150 μm) and high (panels 2: scale bar is 25 μm) magnification images are shown. (a, b) TR146 cells without bacteria. (c, d) Cells infected with S. typhimurium detected with rabbit anti-Salmonella (goat anti-rabbit FITC conjugated). (e, f) Cells infected with L. monocytogenes detected with rabbit anti-Listeria (goat anti-rabbit FITC conjugated). (g) The percentage of cells±s.d., (n=3) in 10 random microscopic fields that mobilized calprotectin. The results represent the average of three independent experiments. (h) TR146 cells with intracellular and extracellular (attached) L. monocytogenes, according to mobilization state. After 2 h invasion, cells were fixed and stained for calprotectin and L. monocytogenes. Mobilized cells (100 counted) and nonmobilized cells (100 counted) were analyzed for presence and location of L. monocytogenes. A significantly (*P=0.046) greater percentage of TR146 cells mobilizing calprotectin had L. monocytogenes located inside the cell, as opposed to nonmobilized cells (mean±s.e., n=3).
In these conditions, invasion by Listeria and Salmonella was confirmed by double immunofluorescence staining. Significantly more cells with intracellular invading Listeria showed mobilized rather than diffuse calprotectin (50±3.3 vs. 33±3.3%, s.e., P<0.05; Figure 2h). Cells frequently showed intra- and extracellular bacteria. Virtually all cells (>90%) showed bound extracellular Listeria, independent of the presence of mobilized calprotectin. When incubated with L. monocytogenes 43249 (MOI of 100) for longer than 2 h, TR146 cells started to die.
Calprotectin colocalizes with microtubules
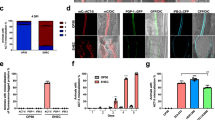
To learn whether mobilized calprotectin associated with cytoskeleton, TR146 cells were incubated with L. monocytogenes 43249 for 2 h at MOI of 100, followed by double immunostaining for calprotectin and β-tubulin. Using confocal analysis (cross-sectional intervals of 0.5 μm), microtubules (anti-β-tubulin fluorescein isothiocyanate (FITC), green) and calprotectin (27E10-biotin-streptavidin Alexa 568, red) showed similar patterns (Figure 3a). When merged, the yellow/orange color strongly suggested colocalization of calprotectin and β-tubulin. In response to L. monocytogenes 43249, calprotectin colocalized with β-tubulin filaments throughout the cell, with more intense fluorescence subjacent to the plasma membrane and in or proximal to the nucleus. Calprotectin and actin did not colocalize.

Colocalization of calprotectin and microtubules. (a) Laser confocal images of TR146 cells (0.5 μm cross-sections through the nucleus) were captured after L. monocytogenes infection. In these nuclear plane images, the microtubule network is shown in green (anti-β-tubulin FITC conjugated) and calprotectin in red (27E10-biotin and streptavidin Alexa 568). When both images were merged, the colocalization between both proteins appears in yellow/orange color. The colocalization was visualized as filaments in the cytoplasm and inside the nucleus. (b) When TR146 cells were incubated with L. monocytogenes (MOI of 100) in the presence of demecolcine 2 μM for 2 h, no filamentous pattern was seen. Fluorescence microscopy of disrupted microtubules is shown in red (anti-β-tubulin and streptavidin Alexa 568), L. monocytogenes in green (detected with rabbit anti-Listeria and goat anti-rabbit FITC conjugated) with DAPI (blue) for nuclear staining. (c) TR146 cells were incubated in the presence of cytochalasin B (2 mM) and then infected with L. monocytogenes (MOI of 100). Actin was stained green with phalloidin conjugated to Oregon Green 514. Calprotectin and Listeria were stained as described for panel b. Scale bars are 25 μm.
TR146 cells were next treated with demecolcine (2 μM for 2 h) to disrupt the microtubule network and then incubated with L. monocytogenes 43249. In demecolcine-treated cells, calprotectin failed to organize into filaments (Figure 3b), whereas Listeria invasion was confirmed by immunofluorescence. As expected, the actin cytoskeleton and cytokeratins were unaffected by demecolcine (data not shown). Phalloidin-stained actin was disrupted by cytochalasin and yet calprotectin mobilized in the presence of Listeria (Figure 3c).
Ca2+ dependency of calprotectin mobilization
To learn whether colocalization of calprotectin with microtubules requires extracellular Ca2+, TR146 cells were incubated with L. monocytogenes 43249 and 2 mM EGTA for 2 h. When extracellular calcium was chelated by EGTA, the frequency of cells with mobilized calprotectin was reduced by 97% when compared to a media control (untreated cells; Figure 4a). Treatment with EGTA did not appear to affect the distribution of microtubules, although the cells appeared more fusiform (data not shown). To determine whether intracellular calcium affects calprotectin mobilization, cells were preincubated for 20 min in glycine, N,N’-(1,2-ethanediylbis(oxy-2,1- phenylene))bis(N-(2-((acetyloxy) methoxy)-2-oxoethyl))-, bis((acetyloxy)methyl) ester (BAPTA/AM; 20 μM) and an additional 20 min in growth media to allow cleavage of the AM groups inside the cell. To release calcium from internal stores, TR146 cells were then incubated with thapsigargin (2 μM, 15 min), washed twice, and incubated with L. monocytogenes 43249 for 2 h. BAPTA/thapsigargin inhibited the frequency of cells showing mobilization by 68% (Figure 4a).

Calprotectin mobilization is Ca2+ dependent, LLO dependent and microtubules abrogate the anti-Listeria effect of calprotectin. (a) TR146 cells were treated with pathway and Ca2+ inhibitors. DMSO was included as a vehicle control. Infection with L. monocytogenes ATCC 43249 occurred at MOI of 100 for 2 h at 37 °C, monolayers were fixed and immunostained with mouse antibody 27E10-biotin to visualize calprotectin as described in the Methods. Cells (500) from five random fields were counted and scored for mobilization. Data represent the percent reduction in frequency of mobilized cells relative to a media control±s.e., n=3. (b) Listeria strains with virulence factor mutations were incubated with TR146 cells at a MOI of 100 for 2 h at 37 °C. Monolayers were fixed and immunostained with mouse antibody 27E10-biotin to visualize calprotectin as described in the Methods. Cells (500) from five random fields were counted and scored for mobilization. Data represent the percentage of cells with mobilized calprotectin±s.e., n=4. (c) Incubation with preformed microtubules inhibits anti-Listeria activity of calprotectin at ED50. Calprotectin at ED50 (250 μg/ml) and tubulin were placed in triplicate wells of microtiter plates at a 1:1 or 0.1:1 molar ratio (tubulin dimer to calprotectin dimer) with a suspension of L. monocytogenes as described in the Experimental Procedures. Plates were incubated for 8 h at 37 °C and bacteria were plated on tryptic-soy agar (TSA) and counted after 24 h. Values are means±s.e. from three separate experiments. At a 1:1 molar ratio, the fold-increase in CFUs in the presence and absence of calprotectin was not significantly different (P>0.05 Duncan's multiple range test). Listeria growth was significantly inhibited in the presence of calprotectin ED50 in all other conditions studied (*P<0.01). Note that no microtubules are present in the vehicle control, DMSO+taxol in tubulin polymerization buffer (TPB).
To learn whether L. monocytogenes-stimulated calcium-signaling pathways18 affect calprotectin mobilization, TR146 cells were preincubated with selected inhibitors for 45 min, washed twice, and then incubated with Listeria. Inhibitors included wortmannin19 and LY294002, inhibitors of phosphatidylinositol 3-kinase,20 U73122, an agonist of phospholipase C,21 and calphostin C, an inhibitor of protein kinase C.22 All inhibited calprotectin mobilization (Figure 4a). Yet in the presence of EGTA, BAPTA/thapsigargin23 or any of the calcium-signaling inhibitors, L. monocytogenes still invaded TR146 cells (data not shown).
Calprotectin mobilization is listeriolysin O-dependent
To determine whether calprotectin mobilization required contact between Listeria and TR146 cells, cells were grown in transwell plates. L. monocytogenes (MOI 100) were placed in the inserts and incubated for 2 h. In this system, TR146 cells were unable to contact Listeria, but were exposed to products, including released LLO. TR146 cells failed to mobilize calprotectin (data not shown). When undiluted medium harvested from L. monocytogenes wild-type strain 10403S was incubated directly with TR146 cells, fewer than 1% of TR146 cells showed mobilized calprotectin.
Several Listeria pathogenicity factors were considered for their contributions to calprotectin mobilization. TR146 cells were incubated with L. monocytogenes 10403S or mutants lacking LLO (DPL2161), LLO, and a broad-range phospholipase C (PC-PLC; DPL2318), or LLO and metalloprotease (DPL2404). Each LLO mutant caused significantly fewer TR146 cells to mobilize calprotectin than the wild-type L. monocytogenes 10403S. In contrast, a L. monocytogenes 10403S mutant lacking ActA (DPL3078) did not significantly affect calprotectin mobilization (Figure 4b).
Polymerized tubulin (microtubules) abrogates the anti-Listeria effect of calprotectin
To determine whether colocalization with microtubules reduces the anti-Listeria effect of calprotectin, we determined the ED50 of purified calprotectin in vitro. Purified calprotectin inhibited growth of L. monocytogenes 43249 at concentrations ranging from 200 to 270 μg/ml. Calprotectin at 250 μg/ml reduced growth by approximately 53% (average of five experiments; ED50). At the ED50, calprotectin inhibition of L. monocytogenes growth was unaffected by 20 μM taxol in dimethyl sulfoxide (DMSO; Figure 4c), which maintained the polymerization state of preformed microtubules in vitro. Taxol itself inhibited Listeria, but this effect was independent and not additive with calprotectin. In the presence of preformed microtubules (1:1 ratio of microtubules to calprotectin complex), inhibition of Listeria was not statistically different in the presence or absence of calprotectin (P>0.05; Duncan's multiple range test). Calprotectin (and taxol) was largely neutralized by preformed microtubules. At a ratio of microtubules to calprotectin of 1:10, Listeria growth was significantly inhibited by calprotectin (P<0.01).
Discussion
In human infections, Listeria and Salmonella in contaminated food and water generally enter the body through the mouth. Remarkably, oral infections by these organisms are unknown. Although contaminated food is macerated and partially retained in proximity to the oral mucosa for minutes to hours, actual infections occur in the gastro-intestinal mucosa. The healthy oral epithelium expresses the antimicrobial protein, calprotectin, which is not found in the intestinal epithelial cells in the absence of inflammation. Hence, one response of the gut epithelium during inflammation24, 25 and to certain enteric pathogens is the upregulation of calprotectin. Calprotectin shows a broad scope of antimicrobial activity against bacteria and fungi10, 12 and may contribute to innate host defense by epithelia.26 In rodent models, enteric infection with Listeria27 or Salmonella28, 29 increases the expression of calprotectin subunits in the intestinal mucosa. Hence, enteric pathogens colonize the healthy intestinal tissues when the intestinal epithelium does not express calprotectin, but as the levels of pathogen increase in the intestines, calprotectin expression is induced in response.
We modeled the effect of calprotectin on invading Listeria and Salmonella in vitro. We show for the first time that calprotectin effectively inhibited growth of Listeria in vitro and within keratinocytes. In the gastrointestinal tract, the spectrum of antimicrobial peptides effective against Listeria also include cathelin-related antimicrobial peptide, which is expressed in the intestinal crypts of newborns,30 Paneth cell mature defensins (cryptdins) and lysozyme,31 and epithelial defensins.32 Calprotectin, however, differs from the other prominent antimicrobial peptides and lysozyme. In the intestinal epithelium, calprotectin both is inducible and expressed in the cytoplasm. Consistent with expression in the cytoplasm, we show that keratinocyte calprotectin inhibits low levels of Listeria invasion and intracellular growth. After invasion, Listeria resides in the cytoplasm. In general, intracellular growth of Salmonella at MOIs of 1, 10, and 100 appears unaffected by calprotectin (data not shown), reflecting that this organism is generally confined to vacuolar or endosomal compartments. At higher MOIs, Listeria induces mobilization of calprotectin in a greater percentage of cells. Mobilized or complexed calprotectin is less effective as an intracellular antimicrobial. Consequently, with greater invasion, Listeria overcomes calprotectin and grows well in the cytoplasm. In the gut, calprotectin will be induced in response to infectious agents in ingested foods. If calprotectin is induced rapidly, epithelial cells may suppress invasion by Listeria. As Listeria proliferates, however, our data suggest that the protective effect of calprotectin could be overcome.
The intracellular activity of calprotectin is independent of cell background. TR146 buccal carcinoma cells33 facilitated study of calprotectin expressed under control of native endogenous promoters. TR146 cells constitutively express calprotectin in their cytoplasm,4 similar to normal mucosal keratinocytes.2 Probed with mAb 27E10, calprotectin distributes diffusely in the cytoplasm, concentrating in the perinuclear area as described previously,4 and the data suggest that cells with diffuse expression of calprotectin show fewer intracellular Listeria than cells with mobilized calprotectin. Like TR146 cells, calprotectin-expressing KB carcinoma cells (HeLa-like), which were transfected to express both subunits of calprotectin under control of exogenous promoters,13 also showed intracellular anti-Listeria activity. The comparison between cell lines and control of expression strongly suggests that calprotectin is the basis for intracellular anti-Listeria activity and not another idiosyncratic feature of the cell.
As the input of Listeria increases to an MOI of 100, the proportion of keratinocytes showing mobilized calprotectin—change from diffuse to filamentous intracellular distribution—increases to about 10% at the time of observation. Mobilization was characterized by LLO-dependent colocalization of calprotectin with microtubules. When colocalized with microtubules, we show that the intracellular anti-Listeria activity of calprotectin was neutralized or subverted. When mixed with equimolar preformed microtubules in vitro, calprotectin-dependent anti-Listeria activity was lost. In the presence of excess calprotectin, anti-Listeria activity was rescued. Anti-Listeria activity appears to require free diffusible calprotectin within cells. Despite the increase in expression of calprotectin in response to enteric infection in the gut, Listeria appears to have evolved an LLO-dependent mechanism to overcome autonomous anti-Listeria immunity in the epithelial cells. Definitive proof in vivo needs to be established.
In TR146 cells, Salmonella and invasive L. monocytogenes LLO-deficient mutants failed to induce significant mobilization of calprotectin. Calcium fluxes and signaling directed by Salmonella effector proteins from within the vacuole or extracellularly34 were also insufficient to trigger calprotectin mobilization. To explain mobilization of calprotectin in response to Listeria, soluble, and cell-surface LLO was considered a candidate agonist, modulating signaling, host cell gene expression, and the formation of Ca2+-permeable pores leading to intracellular Ca2+ oscillations.35 Listeria failed to mobilize calprotectin when separated by transwells or when freshly harvested LLO-containing culture medium was incubated directly with TR146 cells. TR146 cell localization of calprotectin appears unresponsive, therefore, to extracellular soluble LLO, although Listeria at the plasma membrane interface may express higher concentrations of active LLO than we obtained in culture media or across transwell membranes.
Mobilization of calprotectin may depend on the availability of extracellular and intracellular stores of calcium. LLO signals for Ca2+ influx from extracellular reservoirs and release from intracellular stores.35 In response to Listeria, mobilization required both extracellular and intracellular calcium. Keratinocytes were unable to mobilize calprotectin when incubated with EGTA to chelate extracellular calcium or BAPTA/thapsigargin to deplete and chelate intracellular calcium stores. If Listeria signals for mobilization at the cell membrane, resulting phosphotidylinositol hydrolysis mediates intracellular calcium transients and waves.35 Both LLO activity and internalin B-stimulated pathways can generate lipid mediators, which open intracellular calcium channels.18, 35 Although lipid mediators can also be generated intracellularly by Listeria, we show that inhibition of the inositol 1,4,5-triphosphate-dependent calcium-signaling pathway attenuates mobilization of calprotectin. Other downstream inhibitors of internalin B-signaling and other calcium release pathways, wortmannin, LY294002, U73122, Calphostin C, BAPTA/AM, and thapsigargin inhibit downstream Ca2+ release from intracellular stores18, 36 and inhibited the mobilization of calprotectin.
Pore-forming LLO also mediates rupture of the primary vacuole or endosome, allowing entry of Listeria into the cytoplasm.37 As Listeria-invaded TR146 cells most frequently show mobilized calprotectin, LLO-mediated release of intracellular calcium from membrane stores38 may mediate mobilization. Listeria in the cytoplasm also secretes LLO, which is degraded by an ubitquitin-dependent N-end rule pathway.37 In addition to release of calcium from intracellular stores, Listeria are strongly suggested to induce signaling responses mediated by membrane-associated, cytoplasmic pathogen sensors, such as the nucleotide-binding oligomerization domain-leucine-rich repeat.39 Cytosolic signaling through the nucleotide-binding oligomerization domain-leucine-rich repeat may occur during LLO-dependent escape from the endosome,39 resulting in expression of the “late” cluster of IFN-responsive genes.40 Mobilization of calprotectin and colocalization with β-tubulin could, therefore, involve LLO-dependent cytoplasmic pathogen sensor signaling and Listeria-induced release of calcium from intracellular stores.
The availability of intracellular calcium affects the structure and function of calprotectin. As the EF-hands bind calcium,41 calprotectin complexes with specific effector molecules, including arachidonic acid,42 intermediate filaments,1, 4 keratin filaments9 and β-tubulin.43 With elevated intracellular calcium, the calprotectin heterodimer could be expected to form tetramers and higher order multimers.44 Tetramer formation has been associated with ability of calprotectin to promote formation of tubulin microtubules.44 It is unknown, however, whether intracellular anti-Listeria activity and mobilization of calprotectin depend on tetramer formation. Indeed, the oligomerization state of calprotectin complex within cells has not been reported. In our study, intracellular calprotectin was detected with the murine anti-human calprotectin complex-specific monoclonal antibody 27E10. This antibody, although specific for complexes and also colocalizes calprotectin with microtubules (Figure 3a), does not appear to distinguish heterodimers from higher order multimers. It is clear, however, that the mobilization of calprotectin heterodimers to complex with β-tubulin is likely mediated by an intracellular calcium flux initiated by LLO. Mobilization and colocalization of calprotectin with microtubules may also be transient, and sensitive to changes in intracellular calcium. Images illustrating cells with mobilized and colocalized calprotectin and tubulin represent only a snapshot in time. The actual frequency of mobilization may be underestimated in our studies.
Information about our cell model is incomplete, however, as we do not know the concentrations of calprotectin in subcellular microdomains and environments. From our immunofluorescence microscopy images, calprotectin complex distributes nonuniformly after incubation with Listeria. The average concentration of calprotectin complex in a resting keratinocyte is estimated to be 1 ng per μg cell cytosol protein, but intracellular concentrations at discrete sites could simulate the ED50 shown in our in vitro experiments. When calprotectin appears mobilized and complexed with microtubules, however, significantly more intracellular Listeria are seen after invasion. Complexing of calprotectin with microtubules appears to subvert intracellular anti-Listeria activity. For the first time, therefore, we show that invasive L. monocytogenes direct calprotectin to colocalize with β-tubulin by signaling through LLO. This mechanism appears to subvert autologous epithelial cell immunity, enabling Listeria to replicate within the cytoplasm and spread from cell-to-cell.
Methods
Strains, cells, and reagents. Bacterial strains (Table 1) were maintained on tryptic-soy agar (Difco BD, Franklin Lakes, NJ) and grown in brain–heart infusion medium (Difco). KB cells (HeLa-like calprotectin-negative, epithelial cell line, ATCC CCL-17) and calprotectin-expressing transfectants were grown and maintained as described previously.45 A calprotectin-positive human head and neck squamous cell carcinoma cell line TR146 (TR146 cells (gift from Dr Reuben Lotan, University of Texas, MD. Anderson Cancer Center, Houston, TX) was cultured in Ham's F-12 medium with L-glutamine (Cellgro Mediatech Inc, Manassas, VA) supplemented with heat inactivated 10% (v/v) fetal bovine serum (Atlanta Biologicals, Lawrenceville, GA) and maintained at 37 °C in a 5% CO2 humidified atmosphere. All reagents were obtained from Sigma Aldrich, St Louis, MO) unless otherwise specified.
Invasion assay. Bacterial invasion into KB transfected cells was determined by an antibiotic protection assay (Elsinghorst, 1994). L. monocytogenes ATCC 43249, ATCC 19111, 10403S, S. enterica serovar Typhimurium (S. typhimurium) ATCC 14028, and SL 1344 (Table 1) were grown in brain–heart infusion media and on tryptic-soy agar (Difco). Listeria were harvested from log phase (optical density at 620 nm of 0.4–0.6) and Salmonella from stationary phase (optical density 1.4) and used to infect KB transfectants. KB transfectants were grown in Eagle's Minimum Essential Medium (Cellgro) supplemented with 10% fetal bovine serum in the absence of G418 sulfate for one passage (4 days) prior to the assay to eliminate residual intracellular antibiotics. KB transfectants (1.2×105 cells) were seeded in each well of 24-well plate the day before the assay. Bacteria were resuspended in tissue culture media and added to the monolayers at a MOI of 1, 10, or 100 bacteria per eukaryotic cell (i.e., MOI 100=107 colony-forming units (CFUs) of bacteria inoculated onto 105 epithelial cells). Infected monolayers were incubated for 2 h at 37 °C in a 5% CO2–95% air atmosphere. Following this invasion period, the monolayers were washed twice with Dulbecco's phosphate-buffered saline with Ca2+ and Mg2+ and incubated for another 1.5 h in tissue culture media containing gentamicin (100 μg/ml) to kill extracellular bacteria. For time course analyses, the monolayers were further incubated for another 2–4 h in the presence of 20 μg/ml gentamicin. The concentration of gentamicin was reduced during prolonged incubation to minimize uptake into KB cells. After gentamicin incubation, the infected monolayers were washed twice as described above and lysed with distilled water for 15 min at room temperature. Released intracellular bacteria were diluted and plated with a spiral plater (Spiral Biotech, Bethesda, MD), incubated at 37 °C in air, and enumerated by colony count after 24 h. Each invasion assay was performed in triplicate wells and repeated in at least three independent experiments. Control studies were conducted to verify that 100 μg/ml of gentamicin killed 100% of Listeria and Salmonella after 1 h of exposure. The viability of KB transfectants was unaffected by bacterial invasion as determined by trypan blue dye exclusion. Intracellular growth was reported as the fold-increase in CFUs relative to the number of CFUs at 3.5 h post-inoculation as described by Mills and Finlay.46
Bacterial invasion of TR146 cells and incubation with inhibitors. For invasion, TR146 cells (2×105) were grown overnight in six-well plates (Costar Corning Inc. Life Sciences, Lowell, MA) on glass coverslips coated with 0.2% (v/v) gelatin. L. monocytogenes was harvested in mid-logarithmic phase and S. typhimurium from stationary phase as above. Bacteria were added to cell monolayers at an MOI of 100 in 1 ml, and incubated at 37 °C in a 5% CO2–95% air atmosphere for 2 h before immunostaining (described below).
Microtubules were disrupted with demecolcine (2 μM), actin filaments were disrupted with cytochalasin B (2 mM), and to chelate extracellular calcium, ethylene glycol-bis (β-aminoethyl ether)-N,N,N’,N’-tetraacetic acid (EGTA; 2 mM) was used. Each reagent was added to the TR146 monolayer together with L. monocytogenes and incubated for 2 h before immunostaining (see below).
For BAPTA/thapsigargin treatment, incubation for 20 min in BAPTA/AM (Molecular Probes, Invitrogen Corp, Carlsbad, CA, 20 μM) was followed by an additional 20 min incubation in Ham's with 10% fetal bovine serum to allow cleavage of the AM group inside the cell. Thapsigargin (2 μM) was added for 15 min. In separate experiments, TR146 monolayers were treated for 45 min before invasion with wortmannin (100 nM), LY294002 (100 μM), U73122 (2.5 μM), or calphostin C (Calbiochem, San Diego, CA, 2 μM). Cells were washed twice with Dulbecco's phosphate-buffered saline after treatment with BAPTA/thapsigargin, wortmannin, LY294002, U73122, and calphostin C because it was unknown how these inhibitors would affect Listeria during invasion. Monolayers were then incubated with L. monocytogenes for 2 h before immunostaining.
Invasion in each experimental condition was verified by double immunofluorescence staining of intracellular and extracellular L. monocytogenes as described previously13 (described below).
Antibodies. To identify cytoplasmic calprotectin (S100A8/A9 complex), the murine anti-human mAb 27E10-biotin (dilution 1:50; Bachem, Torrance, CA) that does not bind monomeric or homopolymeric subunits17 used. L. monocytogenes was identified with rabbit anti-L. monocytogenes (dilution 1:3,000; Biodesign, Meridian Life Sciences, Saco, ME) and S. typhimurium was identified with rabbit anti-Salmonella (dilution 1:3,000; Biodesign). Microtubules were detected with mouse mAb anti-β-tubulin clone Tub 2.1 (dilution 1:300, Sigma-Aldrich, St Louis, MO) or mouse mAb anti-β-tubulin conjugated with FITC(dilution 1:30, Sigma). Actin filaments were detected by phalloidin conjugated with Oregon Green 514 (dilution 1:50, Molecular Probes). Cytokeratins were detected by mouse mAb anti-pan-cytokeratin clone C-11 (dilution 1:40, Sigma-Aldrich). All monoclonal antibodies were isotype IgG1, and, therefore, the controls used were mAb mouse IgG1 (Sigma) conjugated or not with biotin and polyclonal rabbit IgG (Sigma) were employed. Streptavidin conjugated with Alexa Fluor 568 (dilution 1:2,000; Molecular Probes) or Alexa Fluor 350 (dilution 1:350, Molecular Probes) was used to detect biotinylated antibodies. To detect unlabeled or underivatized primary antibodies, goat anti-rabbit IgG conjugated with FITC (dilution 1:300; Jackson ImmunoResearch, West Grove, PA) or Alexa Fluor 568 (dilution 1:2,000; Molecular Probes) and goat anti-mouse IgG conjugated with Alexa Fluor 568 (dilution 1:2,000, Molecular Probes) were used. The nuclear stain, 4’,6’-diamidino-2-phenylindole dihydrochloride (0.8 mg/ml; Molecular Probes) diluted 1:3,000, was also used. All antibodies and dyes were diluted in PBS with 3% bovine serum albumin.
Immunostaining. After incubation with bacteria, cells were washed twice with Dulbecco's phosphate-buffered saline and fixed for 10 min with 4% paraformaldehyde in PBS. The cells were then washed twice in PBS and permeabilized with 0.2%. Triton X-100 in PBS for 2 min, washed again, and incubated with primary antibody for 1 h at room temperature. The cells were subsequently washed three times and incubated for 1 h at room temperature in the dark with secondary antibody or streptavidin labeled with a fluorescence probe. To verify antibody specificity, the primary antibodies were replaced with isotype-specific IgG. To control for nonspecific binding of secondary antibody, primary antibodies were omitted. After staining, the cover slips were washed three times and mounted on slides with Fluoromount-G (Southern Biotechnology, Birmingham, AL). The edges of coverslips were sealed with nail polish. The slides were examined with a Nikon Eclipse epifluorescence microscope and photographed using a Spot Camera (Diagnostic Instruments Inc.).
Mobilization was reported as the mean of three independent experiments or as noted. In most experiments, cells (500) from five random fields were counted and scored for mobilization by two independent observers.
Double immunofluorescence staining of intracellular and extracellular L. monocytogenes was carried out as described previously.13 Briefly L. monocytogenes adhering to the cell monolayer (extracellular) were stained with rabbit anti-Listeria antibodies for 1 h, washed three times and incubated with goat anti-rabbit antibodies conjugated with Alexa Fluor 568 for 1 h. Cell monolayers were then washed and permeabilized with 0.2% Triton X100 in PBS to allow for staining of both intracellular and extracellular bacteria. Monolayers were incubated with rabbit anti-Listeria antibody for 1 h and then washed and incubated with goat anti-rabbit IgG conjugated with FITC for 1 h. This procedure enabled us to overlay the fluorescent stains and distinguish extracellular bacteria, which were labeled with both fluorescent antibodies, from intracellular bacteria, which were labeled with only one fluorescent antibody. To verify the mobilization state of invaded cells, 27E10-biotin was added with rabbit anti-Listeria antibody after permeabilization and detected with streptavidin, Alexa Fluor® 350 conjugate.
Bacteria were counted inside and outside cells, including 100 cells with mobilized calprotectin and 100 without. In three separate experiments, the mean counts were virtually the same for both operators. Statistical significance was analyzed by repeated-measures analysis of variance (see below).
Confocal images were obtained at cross-sectional intervals of 0.5 μM using a confocal laser microscope (Sharp MRC-1024). All images were further processed and pseudo-colored with Confocal Assistant®4.02 and Adobe®Photoshop® software.
Anti- Listeria activity of calprotectin incubated with preformed microtubules. Purified (>99%) recombinant calprotectin was prepared as described previously,41 concentrated to 10 mg/ml and dialyzed into tubulin polymerization buffer (20 mM 2-N-(morpho-lino)ethanesulfonic acid/K+, 5 mM MgCl2, 100 mM glutamate, 3.4 M glycerol, 60 μM CaCl2, pH 6.8;44 for 8 h with two changes of buffer.
For growth inhibition experiments, overnight cultures of L. monocytogenes were pelleted, resuspended in fresh brain–heart infusion broth, adjusted to a McFarland equivalence of 1.0, and diluted 1:50,000. Lyophilized preformed microtubules (Cytoskeleton, Denver, CO) were reconstituted to 5 mg/ml in tubulin polymerization buffer with 1 mM GTP, containing 20 μM taxol in DMSO (Cytoskeleton). Calprotectin (5 μl) and tubulin were placed in triplicate wells of microtiter plates at a 1:1 or 1:10 molar ratio (tubulin dimer to calprotectin dimer), and 30 μl of diluted L. monocytogenes suspension was added for a final volume of 100 μl. Vehicle control wells (with no microtubules) contained either DMSO or DMSO with taxol or buffer alone (tubulin polymerization buffer with 1 mM GTP). In separate wells, L. monocytogenes was added to buffer alone and immediately plated to determine the initial inoculum. Plates were sealed and incubated for 8 h at 37 °C in an air atmosphere. The contents of each well were then mixed well, diluted, and a portion was plated onto tryptic-soy agar plates for enumeration of CFUs.
Statistical analysis. Analysis of Listeria invasion (Figures 1 and 2e) used repeated-measures analysis of variance where the “subject” (random effect) was “date” and the fixed effect was the independent variable relevant to the experiment. The threshold of significance was P=0.05. for the analysis of variance F-tests. For independent variables with more than two values post hoc tests used the Bonferroni-corrected significance threshold, namely 0.05 divided by the relevant number of comparisons. Standard error (s.e.) was computed using pooled estimates of components of variation. Experiments regarding interactions between calprotectin and tubulin (Figure 4c) were analyzed by a two-way analysis of variance, with the presence or absence of calprotectin as the first factor, and the presence or absence of tubulin as the second factor (buffer alone, DMSO alone, and taxol alone also were groups in that factor). Comparisons between specific pairs of groups were made by Duncan's multiple range test with an experiment-wise α=0.05.
Disclosure
The authors have no conflict of interest to declare.
References
Nacken, W., Roth, J., Sorg, C. & Kerkhoff, C. S100A9/S100A8: myeloid representatives of the S100 protein family as prominent players in innate immunity. Microsc. Res. Tech. 60, 569–580 (2003).
Ross, K.F. & Herzberg, M.C. Calprotectin expression by gingival epithelial cells. Infect. Immun. 69, 3248–3254 (2001).
Donato, R. S100: a multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. Int. J. Biochem. Cell Biol. 33, 637–668 (2001).
Goebeler, M., Roth, J., van den Bos, C., Ader, G. & Sorg, C. Increase of calcium levels in epithelial cells induces translocation of calcium-binding proteins migration inhibitory factor-related protein 8 (MRP8) and MRP14 to keratin intermediate filaments. Biochem. J. 309, 419–424 (1995).
Rammes, A. et al. Myeloid-related protein (MRP) 8 and MRP14, calcium-binding proteins of the S100 family, are secreted by activated monocytes via a novel, tubulin-dependent pathway. J. Biol. Chem. 272, 9496–9502 (1997).
Schlegel Gomez, R. et al. Variational expression of functionally different macrophage markers (27E10, 25F9, RM3/1) in normal gingiva and inflammatory periodontal disease. J. Clin. Periodontol 22, 341–346 (1995).
Eversole, L.R., Miyasaki, K.T. & Christensen, R.E. Keratinocyte expression of calprotectin in oral inflammatory mucosal diseases. J. Oral Pathol. Med. 22, 303–307 (1993).
Kunz, M., Roth, J., Sorg, C. & Kolde, G. Epidermal expression of the calcium binding surface antigen 27E10 in inflammatory skin diseases. Arch. Dermatol Res. 284, 386–390 (1992).
Thorey, I.S. et al. The Ca2+−binding proteins S100A8 and S100A9 are encoded by novel injury-regulated genes. J. Biol. Chem. 276, 35818–35825 (2001).
Miyasaki, K.T., Bodeau, A.L., Murthy, A.R. & Lehrer, R.I. In vitro antimicrobial activity of the human neutrophil cytosolic S-100 protein complex, calprotectin, against Capnocytophaga sputigena. J. Dent. Res. 72, 517–523 (1993).
Sohnle, P.G., Collins-Lech, C. & Wiessner, J.H. Antimicrobial activity of an abundant calcium-binding protein in the cytoplasm of human neutrophils. J. Infect. Dis. 163, 187–192 (1991).
Steinbakk, M. et al. Antimicrobial actions of calcium binding leucocyte L1 protein, calprotectin. Lancet 336, 763–765 (1990).
Nisapakultorn, K., Ross, K.F. & Herzberg, M.C. Calprotectin expression inhibits bacterial binding to mucosal epithelial cells. Infect. Immun. 69, 3692–3696 (2001).
Gaillard, J.L., Berche, P., Frehel, C., Gouin, E. & Cossart, P. Entry of L. monocytogenes into cells is mediated by internalin, a repeat protein reminiscent of surface antigens from gram-positive cocci. Cell 65, 1127–1141 (1991).
Cossart, P. & Lecuit, M. Interactions of Listeria monocytogenes with mammalian cells during entry and actin-based movement: bacterial factors, cellular ligands and signaling. EMBO J 17, 3797–3806 (1998).
Garcia-del Portillo, F. & Finlay, B.B. Invasion and intracellular proliferation of Salmonella within non-phagocytic cells. Microbiologia 10, 229–238 (1994).
Bhardwaj, R.S. et al. The calcium-binding proteins MRP8 and MRP14 form a membrane-associated heterodimer in a subset of monocytes/macrophages present in acute but absent in chronic inflammatory lesions. Eur. J. Immunol. 22, 1891–1897 (1992).
Bierne, H. et al. The invasion protein InIB from Listeria monocytogenes activates PLC- gamma1 downstream from PI 3-kinase. Cell Microbiol. 2, 465–476 (2000).
Ui, M., Okada, T., Hazeki, K. & Hazeki, O. Wortmannin as a unique probe for an intracellular signalling protein, phosphoinositide 3-kinase. Trends Biochem. Sci. 20, 303–307 (1995).
Vlahos, C.J., Matter, W.F., Hui, K.Y. & Brown, R.F. A specific inhibitor of phosphatidylinositol 3-kinase, 2-(4-morpholinyl)-8-phenyl-4H-1-benzopyran-4-one (LY294002). J. Biol. Chem. 269, 5241–5248 (1994).
Bleasdale, J.E. et al. Inhibition of phospholipase C dependent processes by U-73, 122. Adv. Prostaglandin Thromboxane Leukot Res. 19, 590–593 (1989).
Kobayashi, E., Nakano, H., Morimoto, M. & Tamaoki, T. Calphostin C (UCN-1028C) a novel microbial compound is a highly potent and specific inhibitor of protein kinase C. Biochem. Biophys. Res. Commun. 159, 548–553 (1989).
Preston, S.F. & Berlin, R.D. An intracellular calcium store regulates protein synthesis in HeLa cells, but it is not the hormone-sensitive store. Cell Calcium 13, 303–312 (1992).
Aadland, E. & Fagerhol, M.K. Faecal calprotectin: a marker of inflammation throughout the intestinal tract. Eur. J. Gastroenterol. Hepatol. 14, 823–825 (2002).
Leach, S.T. et al. Serum and mucosal S100 proteins, calprotectin (S100A8/S100A9) and S100A12, are elevated at diagnosis in children with inflammatory bowel disease. Scand. J. Gastroenterol. 42, 1321–1331 (2007).
Boman, H.G. Peptide antibiotics and their role in innate immunity. Annu. Rev. Immunol. 13, 61–92 (1995).
Lecuit, M., Sonnenburg, J.L., Cossart, P. & Gordon, J.I. Functional genomic studies of the intestinal response to a foodborne enteropathogen in a humanized gnotobiotic mouse model. J. Biol. Chem. 282, 15065–15072 (2007).
Rodenburg, W., Bovee-Oudenhoven, I.M., Kramer, E., van der Meer, R. & Keijer, J. Gene expression response of the rat small intestine following oral Salmonella infection. Physiol. Genomics 30, 123–133 (2007).
Rodenburg, W. et al. Salmonella induces prominent gene expression in the rat colon. BMC. Microbiol. 7, 84 (2007).
Menard, S. et al. Developmental switch of intestinal antimicrobial peptide expression. J. Exp. Med. 205, 183–193 (2008).
Eisenhauer, P.B., Harwig, S.S. & Lehrer, R.I. Cryptdins: antimicrobial defensins of the murine small intestine. Infect. Immun. 60, 3556–3565 (1992).
Porter, E.M., van Dam, E., Valore, E.V. & Ganz, T. Broad-spectrum antimicrobial activity of human intestinal defensin 5. Infect. Immun. 65, 2396–2401 (1997).
Rupniak, H.T. et al. Characteristics of four new human cell lines derived from squamous cell carcinomas of the head and neck. J. Natl. Cancer Inst. 75, 621–635 (1985).
Pace, J., Hayman, M.J. & Galan, J.E. Signal transduction and invasion of epithelial cells by S. typhimurium. Cell 72, 505–514 (1993).
Kayal, S. & Charbit, A. Listeriolysin O: a key protein of Listeria monocytogenes with multiple functions. FEMS. Microbiol. Rev. 30, 514–529 (2006).
Ireton, K. et al. A role for phosphoinositide 3-kinase in bacterial invasion. Science 274, 780–782 (1996).
Schnupf, P. & Portnoy, D.A. Listeriolysin O: a phagosome-specific lysin. Microbes Infect. 9, 1176–1187 (2007).
Gekara, N.O. et al. The multiple mechanisms of Ca2+ signalling by listeriolysin O, the cholesterol-dependent cytolysin of Listeria monocytogenes. Cell Microbiol. 9, 2008–2021 (2007).
McCaffrey, R.L. et al. A specific gene expression program triggered by Gram-positive bacteria in the cytosol. Proc. Natl. Acad. Sci. USA 101, 11386–11391 (2004).
Stockinger, S. et al. IFN regulatory factor 3-dependent induction of type I IFNs by intracellular bacteria is mediated by a TLR- and Nod2-independent mechanism. J. Immunol. 173, 7416–7425 (2004).
Hunter, M.J. & Chazin, W.J. High level expression and dimer characterization of the S100 EF-hand proteins, migration inhibitory factor-related proteins 8 and 14. J. Biol. Chem. 273, 12427–12435 (1998).
Kerkhoff, C., Klempt, M., Kaever, V. & Sorg, C. The two calcium-binding proteins, S100A8 and S100A9, are involved in the metabolism of arachidonic acid in human neutrophils. J. Biol. Chem. 274, 32672–32679 (1999).
Vogl, T. et al. MRP8 and MRP14 control microtubule reorganization during transendothelial migration of phagocytes. Blood 104, 4260–4268 (2004).
Leukert, N. et al. Calcium-dependent tetramer formation of S100A8 and S100A9 is essential for biological activity. J. Mol. Biol. 359, 961–972 (2006).
Nisapakultorn, K., Ross, K.F. & Herzberg, M.C. Calprotectin expression in vitro by oral epithelial cells confers resistance to infection by Porphyromonas gingivalis. Infect. Immun. 69, 4242–4247 (2001).
Mills, S.D. & Finlay, B.B. Comparison of Salmonella typhi and Salmonella typhimurium invasion, intracellular growth and localization in cultured human epithelial cells. Microb. Pathog. 17, 409–423 (1994).
Bishop, D.K. & Hinrichs, D.J. Adoptive transfer of immunity to Listeria monocytogenes. The influence of in vitro stimulation on lymphocyte subset requirements. J. Immunol. 139, 2005–2009 (1987).
Jones, S. & Portnoy, D.A. Characterization of Listeria monocytogenes pathogenesis in a strain expressing perfringolysin O in place of listeriolysin O. Infect. Immun. 62, 5608–5613 (1994).
Marquis, H., Doshi, V. & Portnoy, D.A. The broad-range phospholipase C and a metalloprotease mediate listeriolysin O-independent escape of Listeria monocytogenes from a primary vacuole in human epithelial cells. Infect. Immun. 63, 4531–4534 (1995).
Skoble, J., Portnoy, D.A. & Welch, M.D. Three regions within ActA promote Arp2/3 complex-mediated actin nucleation and Listeria monocytogenes motility. J. Cell Biol. 150, 527–538 (2000).
Acknowledgements
This study was supported by NIH grants RO1DE11831, P30DE09737 and RO1GM62112. Alexandre A Zaia also was supported by FAPESP from Brazil, grant 99/10079-4. Kanokwan Nisapakultorn was supported by a scholarship from The Royal Thai government. We thank James Hodges and Joel Rudney for assistance with the statistical analysis and Roy Curtiss, Daniel Portnoy, and Darren Higgins for supplying bacterial strains and mutants used in this study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zaia, A., Sappington, K., Nisapakultorn, K. et al. Subversion of antimicrobial calprotectin (S100A8/S100A9 complex) in the cytoplasm of TR146 epithelial cells after invasion by Listeria monocytogenes. Mucosal Immunol 2, 43–53 (2009). https://doi.org/10.1038/mi.2008.63
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mi.2008.63
This article is cited by
-
Effect of Hangeshashinto on calprotectin expression in human oral epithelial cells
Odontology (2016)
-
The Diagnostic Accuracy of Fecal Calprotectin During the Investigation of Suspected Pediatric Inflammatory Bowel Disease: A Systematic Review and Meta-Analysis
American Journal of Gastroenterology (2014)
-
IL-1 receptor regulates S100A8/A9-dependent keratinocyte resistance to bacterial invasion
Mucosal Immunology (2012)
-
The Antimicrobial Heterodimer S100A8/S100A9 (Calprotectin) Is Upregulated by Bacterial Flagellin in Human Epidermal Keratinocytes
Journal of Investigative Dermatology (2010)
