
Substantial progress on how modern humans settled and adapted to the Tibetan Plateau has been achieved in the past few years. In particular, genetic evidence suggests that peopling the Plateau was attributed mainly to the neolithic immigration initiated from northern China ~7 kilo years ago (kya).1, 2 Furthermore, a very small proportion of genetic components showed a restricted distribution in the Tibetans and were estimated to be extraordinarily ancient, thus plausibly reflecting an early dispersal event onto the Plateau during the Late Pleistocene.1 This observation is in good agreement with the previous archeological records.3 Recently, a more extensive study, by analyzing archeological materials collected from 53 Neolithic and Bronze sites in northeast Tibetan Plateau, provides solid evidence in support of the Neolithic immigration into Tibet and, furthermore, suggests a permanent settlement on the high areas of the Plateau likely occurred till 3.6 kya.4 Although some inconsistency on dating the entrance exists between archeological and genetic studies, likely introduced by the methodology in time estimation, evidence from both disciplines supports the notion that the Tibetans can trace their origin to the Neolithic dispersal from northern China.1, 2, 3, 4
Since evidence from both mitochondrial DNA (mtDNA)1, 2 and Y chromosome2 also suggests the existence of genetic relic of the Late Pleistocene settlers in the Tibetans, a new question then arises: how did modern humans move onto the Plateau or, alternatively, how were these ancient genetic components introduced into the Tibetans? To provide more insights into this issue, we collected and analyzed mtDNA data of a total of 53 665 Asian individuals from 814 populations (Figure 1a and Supplementary Table S1). Then, we examined the distribution of haplogroup M62b, a representative of the suggested genetic relic of the Late Pleistocene immigrants, via a motif search strategy.5 Our result reveals that this haplogroup shows a restricted distribution in the Tibetans and has age ~24 kya (Figures 1b and c), in concordance with the previous studies.1, 2 In contrast with the Neolithic-originated components that can find their closer sister lineages in the current northern Chinese populations,1, 2 only few M62b mtDNAs are present sporadically in some surrounding populations, which all locate merely at the terminal branch (Figure 1c) and thus are most likely introduced from the Tibetan populations via recent gene flow. Taking into account the observation of M62a (~15–24 kya; Figures 1b and c), the sister clade of M62b, in India,6 it is then plausible that M62b might have been introduced into the Tibetans via a different way from the northeast Tibetan Plateau entrance. Our extensively searching result confirms this and further reveals the presence of M62a in Bangladesh and Tibet (Figure 1c). A phylogenetic tree of haplogroup M62 is reconstructed on the basis of whole mitochondrial genomes. Evidently, the Indian M62a mtDNAs belong to the same clade, M62a1 (defined by variants 11914, 12793 and a reverse variant at 4561), whereas another clade in M62a, M62a2 (defined tentatively by variants 16491, 16487, 16452 and 16189), contains only a single Tibetan individual. This result, together with the specific distribution pattern of M62b, suggests haplogroup M62 most plausibly to be originated or differentiated in the Tibetans at ~30–45 kya (Figures 1b and c). Intriguingly, haplogroup M68 (with an age of ~60–80 kya7), the sister clade of M62, was observed in mainland Southeast Asia,7 strongly arguing for a south origin of haplogroup M62.
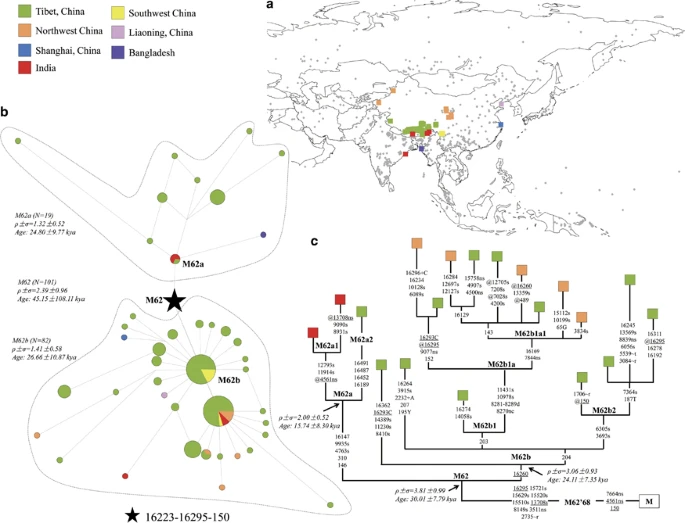
Phylogeographic structure of mtDNA haplogroup M62. (a) Sampling locations. Diamonds in gray indicate the 814 Asian populations (comprising of 53 665 individuals; see Supplementary Table S1), with the one bearing haplogroup M62 being highlighted by square. (b) Median joining network of haplogroup M62. Coalescent ages are calibrated based on mutation rate of control region transitions on segment 16090–1636510 by rho statistic method. (c) Phylogenetic tree of haplogroup M62. The tree is reconstructed by using 19 reported whole mtDNA genomes (see Supplementary Table S2). Nucleotide position numbers are consistent with the revised Cambridge reference sequence (rCRS11). Suffixes C, G and T refer to transversions, ‘d’ means a deletion, ‘s’ means synounymous mutation, ‘ns’ means nonsynounymous mutation and ‘+’ indicates an insertion; recurrent mutations are underlined; ‘@’ means a reverse mutation; and ‘h’ means heterogeneity. Coalescent ages are calibrated based on mutation rate of coding region synonymous mutations10 by rho statistic method. A full color version of this figure is available at the Journal of Human Genetics journal online.
Although more information, especially the complete mtDNA genomes, will be of help to elucidate whether the presence of haplogroup M62 in Tibetans was introduced from South Asian populations6 or in fact originated from the initial settlers who entered East Asia via a southern migration route,8 both scenarios support that M62, different from that of the Neolithic components, has an origin from the south. Consistently, archeological evidence suggests that the foothill of the Himalayas in northeast India serves as an important area for early human movements.9 Meanwhile, it is also suggested that southwestern China was likely the genetic reservoir of the initial settlers, as manifested by an enrichment of mtDNA basal lineages therein.5 These observations add support to the possibility that some additional entrance onto the Tibetan Plateau from the south. Collectively, it becomes evident that peopling the Tibetan Plateau is more complex than expected, likely involving multiple entrance events with different places of origin. More evidence from multiple disciplines, including archeology and genetics, will be of great help to shed more light on this interesting but challenging issue.
References
Zhao, M., Kong, Q. P., Wang, H. W., Peng, M. S., Xie, X. D. & Wang, W. Z. et al. Mitochondrial genome evidence reveals successful late paleolithic settlement on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 106, 21230–21235 (2009).
Qi, X. B., Cui, C. Y., Peng, Y., Zhang, X. M., Yang, Z. H. & Zhong, H. et al. Genetic evidence of Paleolithic colonization and neolithic expansion of modern humans on the Tibetan Plateau. Mol. Biol. Evol. 30, 1761–1778 (2013).
Aldenderfer, M. & Zhang, Y. N. The prehistory of the Tibetan Plateau to the seventh century AD: perspectives and research from China and the West since 1950. J. World. Prehist. 18, 1–55 (2004).
Chen, F. H., Dong, G. H., Zhang, D. J., Liu, X. Y., Jia, X. & An, C. B. et al. Agriculture facilitated permanent human occupation of the Tibetan Plateau after 3600 BP. Science 347, 248–250 (2014).
Kong, Q. P., Sun, C., Wang, H. W., Zhao, M., Wang, W. Z. & Zhong, L. Large-scale mtDNA screening reveals a surprising matrilineal complexity in east asia and its implications to the peopling of the region. Mol. Biol. Evol. 28, 513–522 (2011).
Chandrasekar, A., Kumar, S., Sreenath, J., Sarkar, B., Urade, B. & Mallick, S. et al. Updating phylogeny of mitochondrial DNA macrohaplogroup m in India: dispersal of modern human in South Asian corridor. PLoS ONE 4, e7447 (2009).
Zhang, X. M., Qi, X. B., Yang, Z. H., Serey, B., Sovannary, T. & Bunnath, L. et al. Analysis of mitochondrial genome diversity identifies new and ancient maternal lineages in Cambodian aborigines. Nat. Commun. 4, 2599 (2013).
Macaulay, V., Hill, C., Achilli, A., Rengo, C., Clarke, D. & Meehan, W. et al. Single, rapid coastal settlement of asia revealed by analysis of complete mitochondrial genomes. Science 308, 1034–1036 (2005).
Hazarika, M. Lithic industries with palaeolithic elements in Northeast India. Quatern. Int. 269, 48–58 (2012).
Soares, P., Ermini, L., Thomson, N., Mormina, M., Rito, T. & Rohl, A. Correcting for purifying selection: an improved human mitochondrial molecular clock. Am. J. Hum. Genet. 84, 740–759 (2009).
Andrews, R. M., Kubacka, I., Chinnery, P. F., Lightowlers, R. N., Turnbull, D. M. & Howell, N. Reanalysis and revision of the Cambridge reference sequence for human mitochondrial DNA. Nat. Genet. 23, 147 (1999).
Acknowledgements
This work was supported by grants from the National Natural Science Foundation of China (nos. 31123005, 31322029).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on Journal of Human Genetics website
Supplementary information
Rights and permissions
About this article

Cite this article
Li, YC., Tian, JY. & Kong, QP. A dual origin of Tibetans: evidence from mitochondrial genomes. J Hum Genet 60, 403–404 (2015). https://doi.org/10.1038/jhg.2015.40
Published:
Issue Date:
DOI: https://doi.org/10.1038/jhg.2015.40
