
Abstract
We examined the association of schizophrenia (SCZ) and dihydropyrimidinase-like 2 (DPYSL2), also known as collapsin response mediator protein 2, which regulates axonal growth and branching. We genotyped 20 tag single nucleotide polymorphisms (SNPs) in 1464 patients and 1310 controls. There were two potential associations in a screening population of 384 patients and 384 controls (rs2585458: P=0.046, rs4733048: P=0.014). However, we could not replicate these associations in a confirmatory population of 1080 patients and 926 controls (rs2585458: P=0.39, rs4733048: P=0.70) or a joint analysis (rs2585458: P=0.72, rs4733048: P=0.10). We conclude that DPYSL2 does not have a major function in SCZ in Japanese subjects.
Similar content being viewed by others

Introduction
Schizophrenia (SCZ) is a severe debilitating neuropsychiatric disorder that affects ∼1% of the general population. Family, twin and adoption studies support a substantial genetic contribution to SCZ, but its etiology remains unclear.1 Moreover, irregularities consistent with abnormal brain development, including faulty neuronal migration and altered spatial neuronal arrangement have been reported in SCZ. Together with behavioral, neuromotor and other functional abnormalities that occur in childhood and predict SCZ, such as low IQ, poor motor skills and poor development of language and social skills, these morphological findings indicate a developmental origin for SCZ.2, 3
Dihydropyrimidinase-like 2 (DPYSL2), also known as collapsin response mediator protein 2, is involved in the regulation of axon formation during neuronal polarization.4, 5 Overexpression of Dpysl2 induces the formation of multiple axons and can alter an established dendrite to become an axon, indicating that overexpressed Dpysl2 confers axonal identity not only on immature neurites but also on established dendrites. These observations indicate that DPYSL2 has a crucial function in axon formation of hippocampal neurons, thereby establishing and maintaining neuronal polarity. DPYSL2 interacts with tubulin heterodimers and promotes microtubule assembly in vitro. Thus, DPYSL2 seems to promote neurite elongation and axon specification by regulating microtubule assembly, endocytosis of adhesion molecules and reorganization of actin filaments.6
DPYSL2 is located on chromosome 8p21.2. This region has been reported as positive in meta-analysis of genome-wide linkage studies of SCZ.7 One study showed that chromosomal 8p is a potential hub for developmental neuropsychiatric disorders and contains 21 genes (ADRA1A, ARHGEF10, CHRNA2, CHRNA6, CHRNB3, DKK4, DPYSL2, EGR3, FGF17, FGF20, FGFR1, FZD3, LDL, NAT2, NEF3, NRG1, PCM1, PLAT, PPP3CC, SFRP1 and VMAT1/SLC18A1) that are likely to contribute to the developmental neuropsychiatric disorders (that is SCZ, autism, bipolar disorder and depression) and neurodegenerative disorders (Parkinson's and Alzheimer's disease).8 DPYSL2 is involved in neuropsychiatric disorders’s biology and clearly associated with SCZ and probably with bipolar disorder. The expression of DPYSL2 in the hippocampus is increased in SCZ patients. Expression of the dihydropyrimidinase-related protein 2 in Down's syndrome and Alzheimer's disease brain is downregulated at the mRNA and dysregulated at the protein level. Several clinical studies have described a variety of neurodevelopmental abnormalities in subjects with defects of DPYSL2. DPYSL2 is a marker for escitalopram resistance in stress model of depression. DPYSL2 was reported to be a SCZ susceptibility gene in Japanese subjects.9 However, the results of replication studies using smaller sets of markers have been inconsistent.10, 11 Therefore, to assess whether DPYSL2 has a function in vulnerability to SCZ, we conducted a two-stage case–control association study in a Japanese population.
Materials and methods
Participants
This study was approved by the Ethics Committee of each participating institute, and written informed consent was obtained from each participant. Patients were included in the study if they (1) met DSM-IV criteria for SCZ, (2) were physically healthy and (3) had no mood disorders, substance abuse, neurodevelopmental disorders, epilepsy or known mental retardation. Consensus diagnoses were made by at least two experienced psychiatrists according to DSM-IV criteria on the basis of unstructured interviews with patients and families and review of medical records. The rate of the samples excluded due to the loss of the consensus was <5%. Controls were selected from the general population who had no history of mental disorders based on self-administered questionnaire during sample inclusion step, and based on unstructured diagnostic interview done by a experienced psychiatrist during the blood collection step. All subjects were unrelated to each other, living in the central area of the main land of Japan and self-identified as Japanese population.
Participants consisted of 1464 unrelated Japanese patients with SCZ (age 44.8±15.0 years (mean±s.d.), male 52.6%) and 1310 unrelated healthy controls (age 36.0±13.7 years (mean±s.d.), male 51.0%). In a screening population, participants consisted of 384 unrelated Japanese patients with SCZ (age 50.5±15.1 years (mean±s.d.), male 52.6%) and 384 unrelated healthy controls (age 38.1±14.2 years (mean±s.d.), male 60.3%). In a confirmatory population, participants consisted of 1080 unrelated Japanese patients with SCZ (age 42.7±14.4 years (mean±s.d.), male 49.8%) and 926 unrelated healthy controls (age 35.1±13.36 years (mean±s.d.), male 55.4%). Characterization of general samples and sampling procedures are available elsewhere.12
Genotyping and data analysis
In the screening population of 384 patients and 384 controls, we examined 20 single nucleotide polymorphisms (SNPs) including the positive SNP (rs17666) in an earlier study of a Japanese population.9 The 20 SNPs were selected by Haploview ver4.1 program13 according to the HapMap database (release no. 24, population: Japanese in Tokyo, minor allele frequency >10%). SNP tagging criteria were based on minor allele frequency (>10%) and correlation coefficient (r2) between loci (>0.8) as reported in the HapMap database. We genotyped all SNPs by TaqMan assay (Applied Biosystems Japan, Tokyo, Japan). For quality control, we checked the deviation from the Hardy–Weinberg equilibrium, the sample-wise call rate >95% and the SNP-wise call rate >95%. A power calculation was done by Power Calculator for Genetics Studies.14 We did statistical calculations using PLINK ver2.05.15 The Fisher exact test was used to compare allele frequencies between patients and controls. Significance level was set at P<0.05. As tagging SNPs were selected based on r2, we included imputation as an exploratory method to compute genotypes of SNPs that were not genotyped.16 The starting point of imputation methods is a reference data set such as the HapMap, in which a large set of SNPs is being genotyped. The underlying assumption is that the reference samples, the cases and the controls are all sampled from the same population. Under this assumption, the three populations share the same linkage disequilibrium structure and the same haplotype distribution for every set of SNPs. Thus, the structure of the linkage disequilibrium in the reference population, in conjunction with the structure of the linkage disequilibrium of the observed SNPs within both the cases and the controls, can be used to impute the alleles of a hidden SNP. Imputed SNPs can then be tested for association using an appropriate statistical test. The advantage of imputing untyped SNPs is that the coverage of common variants within the locus of interest can be enhanced, boosting the statistical power and reducing type 2 errors in the screening population. The MACH program was used to calculate the genotypic prediction of 96 untyped SNPs using directly typed information from the 20 SNPs in the screening scan and the HapMap database (recombination map and haplotype data for the Japanese/Chinese population, release 24; phase II).17 The MACH program was recently reported to have similar imputation accuracy rates to IMPUTE and to outperform fastPHASE, PLINK and Beagle.18 The targeted region of imputation was limited to the DPYSL2 locus.
Results
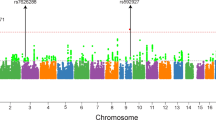
We identified two association signals by PLINK ver2.05 between two SNPs and SCZ before multiple comparisons in the screening population (rs2585458: P=0.046, rs4733048: P=0.014) (Table 1). These two SNPs are located in intron. We imputed the genotype of 96 untyped SNPs using the MACH program and calculated the allelic P-value of each imputed SNP by Haploview ver4.1 (Figure 1). The lowest P-value in imputation was 0.016 at both typed rs4733048 and imputed rs4076071. To confirm the potential associations of these two SNPs and SCZ, we genotyped the two SNPs in a confirmatory population. However, we could not replicate these associations in the confirmatory population of 1080 patients and 926 controls (rs2585458: P=0.39, rs4733048: P=0.70) or in a joint analysis (rs2585458: P=0.72, rs4733048: P=0.10) (Table 2).

Allele P-values of the typed and the imputed SNPs. Red represents typed SNPs and gray represents imputed SNPs. A full color version of this figure is available at the Journal of Human Genetics journal online.
Discussion
In this study, we investigated the association between 20 SNPs of DPYSL2 and SCZ in a Japanese population. We did not observe significant associations between DPYSL2 and SCZ. The positive association observed in the screening population might be due to a type 1 error. After Bonferroni correction, all SNPs in the screening population were negative. Sample size of this study is larger than earlier studies and we used an imputation method, boosting statistic power, so a type 2 error seems unlikely. However, other SNP might be associated with SCZ in the screening sample. Furthermore, we have conducted meta-analysis of data reported previously for rs17666,9, 10, 11 but we could not detect association (fixed model ORci95=0.877–1.175, P=0.843). However, this study may not have sufficient power to detect associations between SNPs with smaller effects and SCZ. It should be noted that a larger sample is required for the detection of a smaller effect. The present sample has statistical power >0.8 for the detection of the role of the polymorphism with a minor allele frequency of 0.1, when the genotype relative risk is 1.35. As all participants in this study were of Japanese descent and recruited from the main island of Japan, the likelihood of population stratification is low.19
There are several limitations in our study. The first limitation in our association study is that cases and controls were not matched in terms of age. In other words, the controls may develop SCZ at some point in life, as they were significantly younger than cases. This might affect the statistic power. The second limitation in our study is that our study design was based on the common disease common variant hypothesis, so we applied minor allele frequency >10% when we selected the 20 tagSNPs. It is difficult to evaluate the association between rare variants and SCZ in our study. In conclusion, this study did not show evidence for the association of DPYSL2 with SCZ in the Japanese population. DPYSL2 may not have a major function in genetic susceptibility to SCZ.
References
Harrison, P. J. & Weinberger, D. R. Schizophrenia genes, gene expression, and neuropathology: on the matter of their convergence. Mol. Psychiatry 10, 40–68 image 45 (2005).
Benes, F. M., Davidson, J. & Bird, E. D. Quantitative cytoarchitectural studies of the cerebral cortex of schizophrenics. Arch. Gen. Psychiatry 43, 31–35 (1986).
Crow, T. J. A. Darwinian approach to the origins of psychosis. Br. J. Psychiatry 167, 12–25 (1995).
Inagaki, N., Chihara, K., Arimura, N., Menager, C., Kawano, Y., Matsuo, N. et al. CRMP-2 induces axons in cultured hippocampal neurons. Nat. Neurosci. 4, 781–782 (2001).
Conde, C. & Caceres, A. Microtubule assembly, organization and dynamics in axons and dendrites. Nat. Rev. Neurosci. 10, 319–332 (2009).
Arimura, N. & Kaibuchi, K. Neuronal polarity: from extracellular signals to intracellular mechanisms. Nat. Rev. Neurosci. 8, 194–205 (2007).
Ng, M. Y., Levinson, D. F., Faraone, S. V., Suarez, B. K., DeLisi, L. E., Arinami, T. et al. Meta-analysis of 32 genome-wide linkage studies of schizophrenia. Mol. Psychiatry 14, 774–785 (2009).
Tabares-Seisdedos, R. & Rubenstein, J. L. Chromosome 8p as a potential hub for developmental neuropsychiatric disorders: implications for schizophrenia, autism and cancer. Mol. Psychiatry 14, 563–589 (2009).
Nakata, K., Ujike, H., Sakai, A., Takaki, M., Imamura, T., Tanaka, Y. et al. The human dihydropyrimidinase-related protein 2 gene on chromosome 8p21 is associated with paranoid-type schizophrenia. Biol. Psychiatry 53, 571–576 (2003).
Hong, L. E., Wonodi, I., Avila, M. T., Buchanan, R. W., McMahon, R. P., Mitchell, B. D. et al. Dihydropyrimidinase-related protein 2 (DRP-2) gene and association to deficit and nondeficit schizophrenia. Am. J. Med. Genet. B. Neuropsychiatr. Genet. 136B, 8–11 (2005).
Zhao, X., Tang, R., Xiao, Z., Shi, Y., Feng, G., Gu, N. et al. An investigation of the dihydropyrimidinase-like 2 (DPYSL2) gene in schizophrenia: genetic association study and expression analysis. Int. J. Neuropsychopharmacol. 9, 705–712 (2006).
Ikeda, M., Takahashi, N., Saito, S., Aleksic, B., Watanabe, Y., Nunokawa, A. et al. Failure to replicate the association between NRG1 and schizophrenia using Japanese large sample. Schizophr. Res. 101, 1–8 (2008).
Barrett, J. C., Fry, B., Maller, J. & Daly, M. J. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics (Oxford, England) 21, 263–265 (2005).
Skol, A. D., Scott, L. J., Abecasis, G. R. & Boehnke, M. Joint analysis is more efficient than replication-based analysis for two-stage genome-wide association studies. Nat. Genet. 38, 209–213 (2006).
Purcell, S., Neale, B., Todd-Brown, K., Thomas, L., Ferreira, M. A., Bender, D. et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 81, 559–575 (2007).
Marchini, J., Howie, B., Myers, S., McVean, G. & Donnelly, P. A new multipoint method for genome-wide association studies by imputation of genotypes. Nat. Genet. 39, 906–913 (2007).
Li, Y., Willer, C., Sanna, S. & Abecasis, G. Genotype imputation. Annu. Rev. Genomics. Hum. Genet. 10, 387–406 (2009).
Pei, Y. F., Li, J., Zhang, L., Papasian, C. J. & Deng, H. W. Analyses and comparison of accuracy of different genotype imputation methods. PloS One 3, e3551 (2008).
Yamaguchi-Kabata, Y., Nakazono, K., Takahashi, A., Saito, S., Hosono, N., Kubo, M. et al. Japanese population structure, based on SNP genotypes from 7003 individuals compared to other ethnic groups: effects on population-based association studies. Am. J. Hum. Genet. 83, 445–456 (2008).
Acknowledgements
We sincerely thank the patients and healthy volunteers for their participation in this study. We express our gratitude to Dr Ryoko Ishihara, Yukako Nakamura, Itaru Kushima, Tomoko Shiino, Eri Yara, Tomo Okochi and Yasuhisa Fukuo for their technical assistance. Funding for this study was provided by research grants from the Ministry of Education, Culture, Sports, Science and Technology of Japan; the Ministry of Health, Labor and Welfare of Japan; Grant-in-Aid for Scientific Research on Pathomechanisms of Brain Disorders from the Ministry of Education, Culture, Sports, Science and Technology of Japan; The Academic Frontier Project for Private Universities, Comparative Cognitive Science Institutes and the Core Research for Evolutional Science and Technology.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Koide, T., Aleksic, B., Ito, Y. et al. A two-stage case–control association study of the dihydropyrimidinase-like 2 gene (DPYSL2) with schizophrenia in Japanese subjects. J Hum Genet 55, 469–472 (2010). https://doi.org/10.1038/jhg.2010.38
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/jhg.2010.38
Keywords
This article is cited by
-
Psychiatric disorders biochemical pathways unraveled by human brain proteomics
European Archives of Psychiatry and Clinical Neuroscience (2017)
-
Evidence for contribution of common genetic variants within chromosome 8p21.2-8p21.1 to restricted and repetitive behaviors in autism spectrum disorders
BMC Genomics (2016)
-
Association study of BCL9 gene polymorphism rs583583 with schizophrenia and negative symptoms in Japanese population
Scientific Reports (2015)
-
CRMPs: critical molecules for neurite morphogenesis and neuropsychiatric diseases
Molecular Psychiatry (2015)
-
Association between DPYSL2 gene polymorphisms and alcohol dependence in Caucasian samples
Journal of Neural Transmission (2014)
