
Abstract
Charcot-Marie-Tooth disease type 1A (CMT1A) and hereditary neuropathy with liability to pressure palsies (HNPP) are caused by a 1.5-Mb duplication and a deletion at chromosome 17p11.2–12 encompassing the peripheral myelin protein 22 gene (PMP22), respectively. We developed a rapid and reliable detection system for duplications/deletions of the PMP22 gene based on measurement of gene copy number. The method involves amplification of a test locus with unknown copy number and a reference locus of known copy number by multiplex PCR (MP), followed by denaturing high-performance liquid chromatography (DHPLC) or capillary electrophoresis detection to identify single copy changes. Thirty-two patients with CMT1A, 17 patients with HNPP, and 61 unaffected individuals were analyzed. Using the same competitive MP protocol, the measured PMP22 gene dosage revealed concordant results between DHPLC and capillary electrophoresis analysis. The results of the MP/DHPLC or the MP/capillary electrophoresis assay were all confirmed by PCR–restriction fragment length polymorphism analysis. We concluded that the MP/DHPLC assay is an efficient, accurate, and reliable technique for gene dosage determination of the PMP22 gene for CMT1A duplication and HNPP deletion. This technique further extends the application of DHPLC as an alternative method for the measurement of gene amplifications and heterozygous deletions in different genetic diseases.
Similar content being viewed by others


Introduction
Although most diseases are caused by point mutations, in many cases, genomic duplications or deletions of genes have also been shown to be responsible for the same disorder. Changes in gene copy number may lead to under- or over-expression of those genes, resulting in a disease phenotype (Lynch 2002). Determination of gene copy number has thus become more important in molecular genetic diagnosis.
Charcot-Marie-Tooth (CMT) disease is the most common inherited disorder of the peripheral nervous system (1/2,500) (Skre 1974). Skre (1974) gives a phenotypic description and a discussion of the genetic heterogeneity of CMT disease type 1, where CMT1A is the most common form of CMT. Around 70% of CMT1A cases are associated with a 1.5-Mb duplication at chromosome 17p11.2–12, where the peripheral myelin protein 22 gene (PMP22) is located (Lupski et al. 1991; Patel et al. 1992; Pentao et al. 1992; Raeymaekers et al. 1991; Timmerman et al. 1992). In contrast, hereditary neuropathy with liability to pressure palsies (HNPP) is caused by a reciprocal deletion of the same region (Chance et al. 1993). In a minority of cases, point mutations in PMP22 can also result in CMT1A and HNPP (Boerkoel et al. 2002).
Many methods to determinate the gene copy number of the PMP22 gene have been reported. Molecular testing by hybridization-based methods includes Southern blotting (Ikegami et al. 1997; MacMillan et al. 1992), pulsed field gel electrophoresis (Lupski et al. 1991; Pentao et al. 1992; Timmerman et al. 1996), and fluorescence in situ hybridization (Lupski et al. 1991; Roa et al. 1996). Other approaches, including analysis of restriction fragment length polymorphism (RFLP) and microsatellite inheritance, have also been commonly used (Badano et al. 2001; Latour et al. 2001; Navon et al. 1995; Roa et al. 1996; Timmerman et al. 1996).
Hybridization-based techniques have several common problems, including the need to use labeled probes and large amounts of DNA together with time-consuming methods. Although microsatellite analysis is a powerful PCR-based technique, its application is limited to cases where parental DNA is available for analysis and where polymorphic markers within the deleted region turn out to be informative in the individual case. Another frequently used method is real-time PCR using standard PCR in conjugation with the fluorescence TaqMan method (Laurendeau et al. 1999; Wilke et al. 2000). However, there are also disadvantages to this approach. Since the results from real-time PCR provide no means of resolving size, the number of loci that can be analyzed in a single tube is currently restricted to four or less by fluorescence profiles and the optical properties of the hardware. Therefore, in applications where multiple loci must be examined, results can only be achieved in multiple tubes, and may be inaccurate due to pipetting errors.
In this study, we introduce a newly developed multiplex PCR (MP)/denaturing high-performance liquid chromatography (DHPLC) assay to determine the gene copy number of PMP22 gene in an efficient way. The assay uses the X-linked CYBB gene and autosomal KRIT1 gene as the standards to determine the relative gene dosage of the PMP22 gene. We demonstrate that this assay is able to accurately distinguish two gene copies from both one copy and three copies; it can thus identify CMT1A patients, HNPP patients, and normal individuals by the accurate determination of PMP22 gene copy number using the DHPLC system.
Materials and methods
Sample preparation
All study subjects were individuals with clinical manifestations of CMT1A/HNPP based on clinical examination and electrophysical diagnosis as carried out previously at National Taiwan University Hospital (Pan et al. 2003; Wilke et al. 2000). Genomic DNA was extracted from peripheral whole blood using the Puregene DNA Isolation Kit (Gentra Systems, Minneapolis, MN) according to the manufacturer’s instructions.
RFLP analysis
All unrelated CMT1A, HNPP patients, and normal control individuals were confirmed by a method previously described (Stronach et al. 1999). The method was a PCR-based test that amplifies a 3.6-kb region including the 1.7-kb hotspot from specific CMT type 1A-repetitions (CMT1A-REPs), allowing the rapid diagnosis of CMT1A and HNPP patients. In the analysis of 32 CMT1A and 17 HNPP patients, duplication and deletion events were detected in all samples with cross-over breakpoints known to be within the region amplified by PCR. Primers were designed using a published sequence proximal to the distal CMT1A-REP sequence (GenBank accession numbers: U41165-distal and U41166-proximal). Primers for the CMT1A test were, CMT1A/HNPP forward (5′-AGGTTGTTTACTCCTTCTTC-3′), and CMT1A reverse (5′-AGATGGAATAGTAGAGCTCAC-3′). Primers for the HNPP test were, CMT1A/HNPP forward (sequence as above), and HNPP reverse (5′-AGATGGAATAGTAGAGTGAG-3′). The amplifications for the CMT1A and HNPP tests were carried out in 50 μl volumes with 1.5 mM MgCl2, 0.4 μM of the appropriate primers, 80 μM dNTPs, 100 ng genomic DNA, 2.6 U Taq DNA polymerase using the Expand Long Template PCR system (Roche Applied Science, Mannheim, Germany). Amplification for CMT1A and HNPP testing comprised an initial denaturation step at 94°C for 3 min followed by ten cycles of 30 s at 94°C, 30 s at 54°C, 3 min at 68°C; 25 cycles of 30 s at 94°C, 30 s at 54°C, 3 min at 72°C, including a 20 s auto-extend function on the extension time to give a final extension of 7 min at 68°C using a multiblock system (MBS) thermocycler (ThermoHybaid, Ashford, UK). Amplified products were digested with EcoRI (New England Biolabs, Beverly, MA) at 37°C for 1 h and electrophoresed at 80 V on 0.8% agarose gels.
Multiplex PCR amplification
Primers for MP amplification were designed to amplify three target sequences, one or two of which lie within the potentially duplicated or deleted target region, the others being reference sequences. Primers were designed to have a similar Tm (the temperature at which 50% of double-stranded DNA is denatured) values and sequence composition to allow the use of similar conditions for the MP system. In the case of quantitation of X chromosome copy number, the CYBB gene was coamplified in the same tube in which quantitation of the X chromosome copy number for males and females was carried out. The KRIT1 gene was another genomic reference and was also coamplified in the same tube to confirm the result.
To amplify the PMP22, CYBB, and KRIT1 genes, primer sequences were as follows: PMP22 exon 3 forward (5′-CTTCTGCTTCTGCTGCCTGT-3′) and reverse (5′-CATTCTGAGGCCACATCCTT-3′), PMP22 exon 4 forward (5′-CTAGGTGGCCAAGATTGGAA-3′) and reverse (5′-GGGATTTTGGGCTAGCTCTT-3′), CYBB exon 13 forward (5′-CGGGAAATTCACCTACTTGC-3′) and reverse (5′-AGCATTATTTGAGCATTTGGC-3′), and KRIT1 exon 8 forward (5′-TTCGAATGGCTACTTCTACCTG-3′) and reverse (5′-AAAACGTCTTTTAAATCAGAGC-3′). The X-linked CYBB gene and KRIT1 gene were used as controls to determine the relative gene dosage of the PMP22 gene. The final volume of the MP was 25 μl containing 100 ng genomic DNA, Set 1 used 0.08 μM of each primer of CYBB and KRIT1 genes, 0.2 μM of the primer of exon 3 of PMP22 gene; Set 2 used 0.08 μM of each primer of CYBB and KRIT1 genes, 0.2 μM of the primer of exon 4 of PMP22 gene; Set 3 used 0.08 μM of the primer of KRIT1 gene, 0.28 μM of the primer of exon 3 and 0.24 μM of the primer of exon 4 of PMP22 gene; 200 μM dNTPs, 0.5 U AmpliTaq Gold enzyme (PE Applied Biosystems, Foster City, CA), and 2.5 μl GeneAmp 10× buffer II (10 mM Tris–HCl, pH 8.3, 50 mM KCl), in 2 mM MgCl2 as provided by the manufacturer. Amplification was performed in an MBS thermocycler (ThermoHybaid). PCR amplification was carried out with an initial denaturation step at 95°C for 10 min, followed by 25 cycles consisting of the denaturation at 94°C for 30 s, the annealing at 56°C for 45 s, the extension at 72°C for 45 s, and a final extension step at 72°C for 10 min.
For capillary electrophoresis analysis, the MP protocol described above with the same primer pairs and condition was applied except that fluorescently labeled primer pairs were used and the cycle number was reduced to 23 cycles.
Capillary electrophoresis and DHPLC analysis
For capillary electrophoresis analysis, 3 μl fluorescently labeled product was mixed well with a mixture containing the Rox-labeled G-500 marker and HiDi Formamide (1:40). This mixture was denatured for 5 min at 95°C. Using a four-color-laser-induced capillary electrophoresis system (ABI Prism 310 Genetic Analyzer), polymer GeneScan POP 4, 1× Genetic Analyzer buffer with EDTA (PE Applied Biosystems), a 47-cm uncoated capillary (Chromatographie Service, Langerwehe, Germany), and sample injection at 15 kV for 5 s, separation of the PCR products was reliably achieved at 15 kV over 26 min.
The DHPLC system used to analyze non-fluorescently labeled PCR products was a Transgenomic Wave Nucleic Acid Fragment Analysis System (Transgenomic, San Jose, CA). DHPLC was carried out on automated HPLC instrumentation equipped with a DNASep column (Transgenomic). The DNASep column contained proprietary 2-mm non-porous alkylated poly(styrenedivinylbenzene) particles. The DNA molecules that elute from the column are detected by scanning with a UV detector at 260 nm. DHPLC grade acetonitrile (9017–03, JT Baker, Phillipsburg, NJ) and triethylammonium acetate (TEAA; Transgenomic, Crewe, UK) were used to constitute the mobile phase. The mobile phases consisted of 0.1M TEAA with 500 μl acetonitrile (eluent A) and 25% acetonitrile in 0.1M TEAA (eluent B). The start- and end-points of the gradient were adjusted according to the size of the PCR products by an algorithm provided by the WAVEmaker system control software (Transgenomic). PCR product (15 μl) was injected for each analysis run. The samples were separated at 50°C. The buffer B gradient increased by 2%/min for 4.5 min at a flow rate of 0.9 ml/min. Generally, the analysis for each injection took about 10 min.
Finally, DHPLC and GeneScan software were used to record peak heights corresponding to the signal from each PCR product.
Data analysis
All MP amplifications were carried out in triplicate for each sample tested. Two target reference PCR product ratios were calculated from each triplicate MP. For known unaffected control samples, the mean of the two ratio values was calculated, and each was scaled to a value of 1.0. All test sample ratios were then scaled accordingly.
To calculate the dosage quotient of the PMP22 gene copy number from unknown samples (U) in comparison with control samples (C), the following formulae were used:
Because the CYBB gene is an X-linked gene, we introduce K(U) and K(C) to represent factors for the unknown and control samples: male=2 and female=1.
Results
In this study, 17 patients with HNPP, 32 patients with CMT1A, and 61 normal controls were analyzed. All diagnoses for duplication or deletion of the PMP22 gene were confirmed by PCR–RFLP analysis (Stronach et al. 1999). Based on the method for capillary electrophoresis analysis of Yau et al. (1996), we could determine the gene dosage of the PMP22 gene from genomic references comprising one gene on an autosomal chromosome (KRIT1 gene, on chromosome 7) and one on the X chromosome (CYBB gene, X-linked). By capillary electrophoresis analysis, we could unambiguously identify all samples with different PMP22 gene copies (Fig. 1).

Use of capillary electrophoresis analysis to identify PMP22 gene copy number. a Electropherogram of the multiplex PCR (MP) in Set 1 (KRIT1/CYBB/exon 3 of PMP22 gene) of DNA samples from the Charcot-Marie-Tooth disease type 1A (CMT1A) (duplication), hereditary neuropathy with liability to pressure palsies (HNPP) (deletion), and normal control. b Scatter plot of capillary electrophoresis results for CMT1A, HNPP, and normal controls. ● CYBB gene (X-linked), ■ KRIT1 gene, ★ exon 3 of PMP22 gene, ⌶ mean ± SD
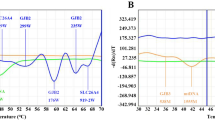
To further explore the possible application of DHPLC to gene dosage analysis of the PMP22 gene, we used the same competitive MP protocol that we had applied previously to the capillary electrophoresis system. In order to determine the optimal PCR cycling number for MP/DHPLC analysis, we ran a series of tests with incremental increases in cycle number (21, 23, 25, 27 cycles). Chromatography using DHPLC of samples with incremental PCR cycle numbers is shown in Fig. 2. All of the results show that the reactions were in linear phase, and 25 cycles was chosen as the number of cycles for quantitative analysis.

Determining PCR cycle number. Using incremental increases in the PCR cycle number from 21 to 27 cycles, the gene ratios for DNA samples from the CMT1A patients (duplication), HNPP patients (deletion), and normal controls are constant
Chromatography using DHPLC analysis of the various combinations of competitive MP is shown in Fig. 3. The MP/DHPLC protocol is similar to that of capillary electrophoresis analysis, and the values for peak height of the autosomal KRIT1 gene and the X-linked CYBB gene were used as genomic references to determine the PMP22 gene copy number. We demonstrated that the dosage of PMP22 gene could be identified correctly by DHPLC analysis, using either the autosomal KRIT1 gene or the X-linked CYBB gene as a genomic reference (Fig. 4). Using both DHPLC and capillary electrophoresis, the measured PMP22 gene copy number was between 1.5 and 2.5 for the normal population, below 1.5 for HNPP patients and above 2.5 for CMT1A patients. In this study, there was no overlapping or ambiguous data observed for either DHPLC or capillary electrophoresis.

Chromatography of denaturing high-performance liquid chromatography (DHPLC) analysis of three sets of DNA samples from two unaffected controls (one male and one female) compared with DNA from two patients with CMT1A (duplication) or with HNPP (deletion). a MP Set 1 (CYBB/KRIT1/exon 3 of PMP22 gene). b MP Set 2 (CYBB/KRIT1/exon 4 of PMP22 gene), c MP Set 3 (KRIT1/exon 3 of PMP22 gene/exon 4 of PMP22 gene). ● CYBB gene (X-linked), ■ KRIT1 gene, ★ exon 3 of PMP22 gene, ▲ exon 4 of PMP22 gene

Scatter plot of DHPLC results for CMT1A, HNPP, and normal controls; the ratio values lie in non-overlapping groups corresponding to the presence of one, two, or three copies of the PMP22 gene, respectively. a Results measured by MP Set 1 (KRIT1/CYBB/exon 3 of PMP22 gene). b Results measured by MP Set 2 (KRIT1/CYBB/exon 4 of PMP22 gene). c Results measured by MP Set 3 (KRIT1/exon 3 of PMP22 gene/exon 4 of PMP22 gene). In a and b, copy numbers are calculated using the CYBB gene as genomic reference in the plots on the left, and using the KRIT1 gene as genomic reference in the plots on the right. In c, the copy numbers are calculated using exon 3 of the PMP22 gene in the left plot and by exon 4 of PMP22 gene in the right plot (KRIT1 gene is as genomic reference). ⌶ Mean ±SD
Table 1 lists the expected and calculated copy numbers of the PMP22 gene as determined using both capillary electrophoresis and DHPLC analysis for the samples, including all the patients with HNPP, CMT1A, and the normal controls. The correlation of the calculated PMP22 gene copy number for all the samples tested between the capillary electrophoresis and DHPLC analysis is shown in Fig. 5, and the correlation coefficient is 0.923 (P<0.01).

Correlation of measured PMP22 gene copy number distribution in the DHPLC system with that in the capillary electrophoresis system
Comparing the time needed for these three methods, PCR–RFLP analysis required 5 h for the long-range PCR and 30 min for enzyme treatment, while capillary electrophoresis required 2 h for the PCR and about 90 min to analyze the data. However, DHPLC was the quickest, requiring 2 h for the PCR and only 30 min for data analysis.
Discussion
Since CMT1A and HNPP are frequent autosomal dominant disorders of inherited peripheral neuropathy, it is important to identify PMP22 gene dosage for diagnostic purpose and genetic counseling. Various methods have been described to quantify gene/exon copy numbers, including microarray hybridization (Zhu et al. 2003), real-time PCR (Kim et al. 2003; Thiel et al. 2003), hybridization with amplifiable probes, and auto-electrophoresis analysis (Akrami et al. 2003; Frisso et al. 2004; Rowland et al. 2001; Yau et al. 1996). Although these methods are reliable, they also carry some limitations when applied to clinical situations. Most of the major problems are time, expense, radioisotope/fluorescence probe labeling, and the need for large amounts of DNA. Hence, we have developed a rapid multiplex competitive PCR-based quantitative technique for relative DNA quantitation (Su et al. 2005).
In this study, the MP approach is a relatively quantitative method. We utilize one or two genomic references to determine the copy number of the gene to be examined: quantitation of the target gene is relative to reference genes of known copy number (Celi et al. 1993; Dehainault et al. 2004; Noonan et al. 1990). A fragment from within a potentially duplicated or deleted target region is amplified simultaneously with a disomic reference region in an MP system. Based on the observation that the amount of PCR product generated from each site of amplification is proportional to the amount of starting template, the ratio of the amount of PCR product generated from each amplification reaction indicates whether there is duplication, deletion, or no change in the target area. Detection of PCR products is carried out either by DHPLC or capillary electrophoresis, both of which provide the sensitivity required for detection of single-copy dosage changes.
A major advantage of this MP approach is that the analysis is carried out within a single tube; therefore, any factors influencing the PCR will affect both reactions in a similar manner and will not alter the final ratio. An additional advantage of the technique is the coupling of DHPLC/capillary electrophoresis and detection in a single system, which is suitable for large-scale sample analysis.
To calculate PMP22 gene dosage by competitive MP strategy, we designed primer pairs for the PMP22 gene to amplify exon 3 and exon 4 in combination, using two other genes (one autosomal KRIT1 gene, one X-linked CYBB gene) as the genomic references. Three independent primer sets with different combinations between these four PCR fragments (KRIT1/CYBB/exon 3 of PMP22 gene; KRIT1/CYBB/exon 4 of PMP22 gene; KRIT1/exon 3 of PMP22 gene/exon 4 of PMP22 gene) were evaluated to test the validity of the assay. These three different sets all yielded concordant and satisfactory results.
The PMP22 gene dosages determined by the MP assay were analyzed by both DHPLC and capillary electrophoresis systems, with concordant results. Capillary electrophoresis is one of the most recent developments in the detection of gene dosage (Beckmann and Schroder 2000; Latour et al. 2001; Poropat and Nicholson 1998; Young et al. 1998). This method is automatic, accurate, and fast; however, it requires fluorescently labeled primer pairs, and the post-processing of PCR products, which requires HiDi formamide for denaturation for 5 min at 95°C, followed by the use of specific sample tubes or plates to carry out the analysis. DHPLC has been used previously to detect point mutations and appears to be an economical and highly sensitive method of mutation detection. Here, we report a new application of this system that is sensitive enough to detect changes in gene copy number from one to two to three copies of the PMP22 gene. Compared with capillary electrophoresis, DHPLC does not require fluorescence labeling of the PCR primers, and unpurified PCR products can be used. This method required only 2 h for PCR and about 30 min of DHPLC to analysis all three MP sets. The average cost of DHPLC analysis is less than US$ 3 per sample. All the above show that this method is very well suited to diagnosing a duplication causing CMT1A or a deletion causing HNPP.
Dehainault et al. (2004) described an application to detect gene rearrangements of the RB1 gene using MP and DHPLC. Their report used the same principles as MP/DHPLC to determine relative gene dosage. They used fluorescence to detect gene dosage, thereby increasing sensitivity by ~100-fold compared with UV detection. In our study, we introduced a reliable method using DHPLC with non-fluorescent detection to quantify PMP22 gene copy number, which is more cost-effective with same resolution. Furthermore, Dehainault et al. (2004) improved the amplification of high GC content DNA using a touchdown protocol. The principles of the touchdown protocol raise a doubt about equitable competition and the results may not be a very precise quantification of gene dosage. In our previous work, we have successfully demonstrated the application of DHPLC to gene dosage analysis of the SMN1/SMN2 genes of spinal muscular atrophy. However, amplification of DNA sequences with high GC content is also a challenge in this study and requires a lot of time to choose the appropriate primer combination for our competitive PCR protocols.
We have demonstrated that MP coupled with DHPLC can accurately distinguish two gene copies from one and three copies, and proved that it is a rapid, reliable, low-cost system for the genetic diagnosis of duplication and deletions. Thus, further exploitation of the precise measurement of gene copy number using this protocol can be expected in the future.
In conclusion, MP/DHPLC analysis permits high sample throughput and can be used for large-scale screening projects. This method is a rapid, reliable, low-cost, and non-fluorescence-based test system for the genetic diagnosis of the CMT1A duplication and the HNPP deletion. It can be used as an alternative to previous methods for detection of these genetic events and can also be extended to the measurement of any copy number defects that are associated with a genetic disease.
References
Akrami SM, Rowland JS, Taylor GR, Armour JA (2003) Diagnosis of gene dosage alterations at the PMP22 gene using MAPH. J Med Genet 40(11):e123
Badano JL, Inoue K, Katsanis N, Lupski JR (2001) New polymorphic short tandem repeats for PCR-based Charcot-Marie-Tooth disease type 1A duplication diagnosis. Clin Chem 47:838–843
Beckmann A, Schroder JM (2000) Screening for Charcot-Marie-Tooth type 1A and hereditary neuropathy with liability to pressure palsy in archival nerve biopsy samples by direct-double-differential PCR. Acta Neuropathol 100:459–463
Boerkoel CF, Takashima H, Garcia CA, Olney RK, Johnson J, Berry K, Russo P, Kennedy S, Teebi AS, Scavina M et al (2002) Charcot-Marie-Tooth disease and related neuropathies: mutation distribution and genotype–phenotype correlation. Ann Neurol 51:190–201
Celi FS, Zenilman ME, Shuldiner AR (1993) A rapid and versatile method to synthesize internal standards for competitive PCR. Nucleic Acids Res 21:1047
Chance PF, Alderson MK, Leppig KA, Lensch MW, Matsunami N, Smith B, Swanson PD, Odelberg SJ, Disteche CM, Bird TD (1993) DNA deletion associated with hereditary neuropathy with liability to pressure palsies. Cell 72:143–151
Dehainault C, Lauge A, Caux-Moncoutier V, Pages-Berhouet S, Doz F, Desjardins L, Couturier J, Gauthier-Villars M, Stoppa-Lyonnet D, Houdayer C (2004) Multiplex PCR/liquid chromatography assay for detection of gene rearrangements: application to RB1 gene. Nucleic Acids Res 32:e139
Frisso G, Carsana A, Tinto N, Calcagno G, Salvatore F, Sacchetti L (2004) Direct detection of exon deletions/duplications in female carriers of and male patients with Duchenne/Becker muscular dystrophy. Clin Chem 50:1435–1438
Ikegami T, Ikeda H, Chance PF, Kiyosawa H, Yamamoto M, Sobue G, Ohnishi A, Tachi N, Hayasaka K (1997) Facilitated diagnosis of CMT1A duplication in chromosome 17p11.2–12: analysis with a CMT1A-REP repeat probe and photostimulated luminescence imaging. Hum Mutat 9:563–566
Kim SW, Lee KS, Jin HS, Lee TM, Koo SK, Lee YJ, Jung SC (2003) Rapid detection of duplication/deletion of the PMP22 gene in patients with Charcot-Marie-Tooth disease type 1A and hereditary neuropathy with liability to pressure palsy by real-time quantitative PCR using SYBR Green I dye. J Korean Med Sci 18:727–732
Latour P, Boutrand L, Levy N, Bernard R, Boyer A, Claustrat F, Chazot G, Boucherat M, Vandenberghe A (2001) Polymorphic short tandem repeats for diagnosis of the Charcot-Marie-Tooth 1A duplication. Clin Chem 47:829–837
Laurendeau I, Bahuau M, Vodovar N, Larramendy C, Olivi M, Bieche I, Vidaud M, Vidaud D (1999) TaqMan PCR-based gene dosage assay for predictive testing in individuals from a cancer family with INK4 locus haploinsufficiency. Clin Chem 45:982–986
Lupski JR, de Oca-Luna RM, Slaugenhaupt S, Pentao L, Guzzetta V, Trask BJ, Saucedo-Cardenas O, Barker DF, Killian JM, Garcia CA et al (1991) DNA duplication associated with Charcot-Marie-Tooth disease type 1A. Cell 66:219–232
Lynch M (2002) Genomics. Gene duplication and evolution. Science 297:945–947
MacMillan JC, Upadhyaya M, Harper PS (1992) Charcot-Marie-Tooth disease type 1a (CMT1a): evidence for trisomy of the region p11.2 of chromosome 17 in south Wales families. J Med Genet 29:12–13
Navon R, Timmerman V, Lofgren A, Liang P, Nelis E, Zeitune M, Van Broeckhoven C (1995) Prenatal diagnosis of Charcot-Marie-Tooth disease type 1A (CMT1A) using molecular genetic techniques. Prenat Diagn 15:633–640
Noonan KE, Beck C, Holzmayer TA, Chin JE, Wunder JS, Andrulis IL, Gazdar AF, Willman CL, Griffith B, Von Hoff DD et al (1990) Quantitative analysis of MDR1 (multidrug resistance) gene expression in human tumors by polymerase chain reaction. Proc Natl Acad Sci USA 87:7160–7164
Pan CL, Tseng TJ, Lin YH, Chiang MC, Lin WM, Hsieh ST (2003) Cutaneous innervation in Guillain-Barre syndrome: pathology and clinical correlations. Brain 126:386–397
Patel PI, Roa BB, Welcher AA, Schoener-Scott R, Trask BJ, Pentao L, Snipes GJ, Garcia CA, Francke U, Shooter EM et al (1992) The gene for the peripheral myelin protein PMP-22 is a candidate for Charcot-Marie-Tooth disease type 1A. Nat Genet 1:159–165
Pentao L, Wise CA, Chinault AC, Patel PI, Lupski JR (1992) Charcot-Marie-Tooth type 1A duplication appears to arise from recombination at repeat sequences flanking the 1.5 Mb monomer unit. Nat Genet 2:292–300
Poropat RA, Nicholson GA (1998) Determination of gene dosage at the PMP22 and androgen receptor loci by quantitative PCR. Clin Chem 44:724–730
Raeymaekers P, Timmerman V, Nelis E, De Jonghe P, Hoogendijk JE, Baas F, Barker DF, Martin JJ, De Visser M, Bolhuis PA, et al (1991) Duplication in chromosome 17p11.2 in Charcot-Marie-Tooth neuropathy type 1a (CMT 1a). The HMSN Collaborative Research Group. Neuromuscul Disord 1:93–97
Roa BB, Greenberg F, Gunaratne P, Sauer CM, Lubinsky MS, Kozma C, Meck JM, Magenis RE, Shaffer LG, Lupski JR (1996) Duplication of the PMP22 gene in 17p partial trisomy patients with Charcot-Marie-Tooth type-1 neuropathy. Hum Genet 97:642–649
Rowland JS, Barton DE, Taylor GR (2001) A comparison of methods for gene dosage analysis in HMSN type 1. J Med Genet 38:90–95
Skre H (1974) Genetic and clinical aspects of Charcot-Marie-Tooth’s disease. Clin Genet 6:98–118
Stronach EA, Clark C, Bell C, Lofgren A, McKay NG, Timmerman V, Van Broeckhoven C, Haites NE (1999) Novel PCR-based diagnostic tools for Charcot-Marie-Tooth type 1A and hereditary neuropathy with liability to pressure palsies. J Peripher Nerv Syst 4:117–122
Su YN, Hung CC, Li H, Lee CN, Cheng WF, Tsao PN, Chang MC, Yu CL, Hsieh WS, Lin WL, et al (2005) Quantitative analysis of SMN1 and SMN2 genes based on DHPLC: a highly efficient and reliable carrier-screening test. Hum Mutat 25:460–467
Thiel CT, Kraus C, Rauch A, Ekici AB, Rautenstrauss B, Reis A (2003) A new quantitative PCR multiplex assay for rapid analysis of chromosome 17p11.2–12 duplications and deletions leading to HMSN/HNPP. Eur J Hum Genet 11:170–178
Timmerman V, Nelis E, Van Hul W, Nieuwenhuijsen BW, Chen KL, Wang S, Ben Othman K, Cullen B, Leach RJ, Hanemann CO et al (1992) The peripheral myelin protein gene PMP-22 is contained within the Charcot-Marie-Tooth disease type 1A duplication. Nat Genet 1:171–175
Timmerman V, Lofgren A, Le Guern E, Liang P, De Jonghe P, Martin JJ, Verhalle D, Robberecht W, Gouider R, Brice A et al (1996) Molecular genetic analysis of the 17p11.2 region in patients with hereditary neuropathy with liability to pressure palsies (HNPP). Hum Genet 97:26–34
Wilke K, Duman B, Horst J (2000) Diagnosis of haploidy and triploidy based on measurement of gene copy number by real-time PCR. Hum Mutat 16:431–436
Yau SC, Bobrow M, Mathew CG, Abbs SJ (1996) Accurate diagnosis of carriers of deletions and duplications in Duchenne/Becker muscular dystrophy by fluorescent dosage analysis. J Med Genet 33:550–558
Young P, Stogbauer F, Wiebusch H, Lofgren A, Timmerman V, Van Broeckhoven C, Ringelstein EB, Assmann G, Funke H (1998) PCR-based strategy for the diagnosis of hereditary neuropathy with liability to pressure palsies and Charcot-Marie-Tooth disease type 1A. Neurology 50:760–763
Zhu D, Kennerson M, Merory J, Chrast R, Verheijen M, Lemke G, Nicholson G (2003) Refined localization of dominant intermediate Charcot-Marie-Tooth neuropathy and exclusion of seven known candidate genes in the region. Neurogenetics 4:179–183
Acknowledgment
This work was supported by Grants from the National Science Council of Taiwan (NSC 93-2314-B-002 174).
Author information
Authors and Affiliations
Corresponding author
Additional information
Chia-Yun Lin and Yi-Ning Su share the first authorship.
Rights and permissions
About this article
Cite this article
Lin, CY., Su, YN., Lee, CN. et al. A rapid and reliable detection system for the analysis of PMP22 gene dosage by MP/DHPLC assay. J Hum Genet 51, 227–235 (2006). https://doi.org/10.1007/s10038-005-0350-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10038-005-0350-9
