
Abstract
In the course of our screening program for inhibitors of hepatic glucose production in rat hepatoma H4IIE-C3 cells, which were used as model liver cells, five naphtoquinone derivatives—javanicin, solaniol, 9-O-methylfusarubin, 5,10-dihydroxy-1,7-dimethoxy-3-methyl-1H-naphtho[2,3-c]pyran-6,9-dione, 9-O-methylbostrycoidin—and vanillin were selected from our natural product library. These naphtoquinone derivatives inhibited hepatic glucose production at IC50 values of 3.8–29 μM, but showed cytotoxicity against hepatic cells after incubation for 48 h. However, vanillin showed an IC50 value of 32 μM without exhibiting cytotoxicity at 50 μM. Therefore, we examined 12 vanillin derivatives to investigate their inhibitory activities against glucose production. Among these analogs, 4-hydro-3-methoxyacetophenone and 5-nitrosalicylaldehyde exhibited stronger inhibition than the other compounds at IC50 values of 25 and 24 μM, respectively, with no cytotoxicity at a concentration of 50 μM. Hence, 4-hydro-3-methoxyacetophenone and 5-nitrosalicylaldehyde may be useful as a lead compound of anti-type 2 diabetic drugs.
Similar content being viewed by others
Main
Type-2 diabetes is a severe disease that is prevalent in the developed world, and the incidence of diabetes is increasing even in developing countries. Type-2 diabetes is associated with impaired glucose clearance by the liver with elevated glucose production in the postprandial state and is characterized by insulin resistance.1 Rigid control of plasma glucose levels is known to decrease the incidence and progression of diabetic complications. The liver maintains blood glucose homeostasis by the absorption of glucose in the postprandial state and by the production of glucose from glycogenolysis and gluconeogenesis in the postabsorptive state. In normal tissues, insulin stimulates glucose uptake by muscles and adipose tissue, and inhibits hepatic glucose production. However, this response to insulin is defective in patients with type-2 diabetes, resulting in long-term hyperglycemia. Therefore, the key to treating diabetes is to reverse or bypass the deficient insulin-signaling pathway in patients with type-2 diabetes. Agents that can inhibit hepatic glucose production through a signaling pathway that is distinct from the insulin-signaling pathway may provide new avenues for inhibiting the elevated glucose production caused by insulin resistance in the case of type-2 diabetes.
Several popular classes of oral therapeutic drugs for diabetes that are available in the market (for example, sulphonylureas and peroxisome proliferator-activated receptor-γ agonists) lower plasma glucose levels by increasing glucose metabolism either through enhanced insulin secretion or improved sensitivity to insulin. Metformin is the only drug that acts primarily by reducing endogenous glucose production without exhibiting a major effect on insulin signaling.2 Metformin lowers glucose levels without resulting in weight gain. Thus, it is often used to treat patients with type-2 diabetes despite the well-known safety concerns in certain diabetic patient populations and the overall high incidence of gastrointestinal intolerance associated with its use. Hence, agents such as metformin are expected to inhibit hepatic glucose production. We screened inhibitors from our natural product library for their effects on glucose production in the rat liver cell line H4IIE-C3 and selected five naphtoquinone derivatives and vanillin as inhibitors. Further, we also evaluated commercially available and synthesized vanillin derivatives for their inhibitory effects on glucose production.
Materials and methods
General experimental procedures
UV and IR spectra were measured on a DU730 spectrophotometer (Beckman Coulter, Fullerton, CA, USA) and an FT-720 spectrophotometer (Horiba, Kyoto, Japan), respectively. High-resolution electrospray ionization-mass spectrometric (HR-ESI-MS) data were recorded on an LCT-Premier XE mass spectrometer (Waters, Milford, MA, USA). NMR spectra were measured on an NMR System 500 NB CL (Varian, Palo Alto, CA, USA) in CDCl3 with the residual solvent peak as an internal standard (δH 7.24 p.p.m.). Normal-phase medium-pressure liquid chromatography (MPLC) was performed using a Purif-pack SI-60 column (Moritex, Tokyo, Japan). Analytical reverse-phase HPLC was performed on an L-column2 ODS column (4.6 i.d. × 150 mm, Chemical Evaluation and Research Institute, Tokyo, Japan) with a 2996 photodiode array detector (Waters) and a 3100 mass detector (Waters). Preparative reverse-phase HPLC was performed on an L-column2 ODS (20 i.d. × 150 mm) with an L-2455 photodiode array detector (Hitachi High Technologies, Tokyo, Japan). The reagents and solvents used in our study were of the highest grade available.
Cell culture
Rat hepatoma cell line H4IIE-C3 was used in this study. The cells were grown at 37 °C in an atmosphere containing 5% CO2 in Minimum Essential Medium Eagle (Sigma, Saint Louis, MO, USA). This medium consisted of 10% fetal bovine serum (Sigma), a 100-μM solution of non-essential amino acid (Invitrogen, Carlsbad, CA, USA), 1 mM sodium pyruvate solution (Invitrogen), supplemented with penicillin (100 U ml−1) and streptomycin (100 μg ml−1).
Assay of glucose production in hepatic cells
For performing the screening, H4IIE-C3 cells were inoculated onto a 384-well plate (Cat. No.781182, Greiner Bio-one, Frickenhausen, Germany) at a density of 1.2 × 104 cells per well and incubated overnight in the serum-containing medium. They were then transferred to a serum-free medium and incubated overnight. Next, the medium was replaced with a glucose-free medium—20 μl of Dulbecco's Modified Eagle's Base—which did not contain L-glutamine, glucose, phenol red, sodium pyruvate and sodium bicarbonate (Sigma), supplemented with 3.7 mg l−1 sodium bicarbonate and 20 mM pyruvic acid. The samples (0.2 μl) to be screened were then added to the medium by using Multi-dispenser ADS-384–8 (BioTec, Tokyo, Japan) and incubated for 5 h. Subsequently, 10 μl of the medium was transferred to 384-well black plates (Cat. No.784900, Greiner) by using ADS-384–8, and the amount of glucose accumulated in the wells was detected by mixing the contents of the well with 10 μl of Amplex Red Glucose Assay kit solution (Invitrogen).
To confirm the activities of the hit samples, the cells were inoculated into a 96-well plate (Cat. No.353072; BD Biosciences, San Jose, CA, USA) at a density of 6 × 104 cells per 100 μl for each well and incubated as described above. After treatment with the hit samples at various concentrations, 40 μl of the medium was transferred to a 96-well black plate (Cat. No. 3916; Corning Inc., Corning, NY, USA), and the amount of glucose accumulated was determined by mixing 40 μl of the Amplex Red Glucose Assay kit solution.
Cytotoxicity assay
Cytotoxic activities were evaluated by WST-8 (2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt) colorimetric assay. The 96-well plate was inoculated with H4IIE-C3 cells at a density of 1 × 104 cells per well; these were subsequently treated with compounds at various concentrations and incubated for 48 h. Next, 10 μl of the WST-8 reagent (Cell Counting Kit; Dojindo, Kumamoto, Japan) was added and the cells were incubated for 1 h at 37 °C in a humidified atmosphere containing 5% CO2. Absorbance of the formazan dye formed was measured at 450 nm.
Natural product library
Our in-house natural product library was used as the source of samples for screening. The library contained 40 508 diverse samples of crude metabolites of actinomycetes (20 608 samples), bacteria (6336 samples) other than actinomycetes and fungi (12 672 samples). It also included plant extracts (252 samples) and metabolites isolated from microorganisms (640 samples). All the samples were dissolved in dimethyl sulfoxide.
Compounds
Javanicin3 (1) and solaniol4 (2) were purified from the culture of Fusarium sp. no. 351386, which was isolated from a soil sample collected from Tanzawa, Kanagawa Prefecture, Japan, with MPLC and HPLC by activity-guided separation.
9-O-Methylfusarubin5 (3), 5,10-dihydroxy-1,7-dimethoxy-3-methyl-1H-naphtho[2,3-c]pyran-6,9-dione6 (4), 9-O-methylbostrycoidin5 (5) and vanillin (6) were selected as hit samples from our purified natural product library. These samples were of fungal origin.
Isovanillin (3-hydroxy-4-methoxybenzaldehyde) (8), 4-hydroxy-3-methoxyacetophenone (9), 4-hydroxy-3,5-dimethoxyacetophenone (10), 5-chlorosalicylaldehyde (12), 5-nitrosalicylaldehyde (13) and 2,4-dihydroxybenzaldehyde (14) were purchased from Tokyo Chemical Industry (Tokyo, Japan); vanillic acid (7) and 5-bromosalicylaldehyde (11) were purchased from Wako Pure Chemical (Osaka, Japan); 2,4,6-trihydroxybenzaldehyde (15) was purchased from Aldrich (Saint Louis, MO, USA).
Hydroxy-4,6-dimethoxybenzaldehyde (16)
Oxalyl chloride (16.0 ml, 183 mmol, 2.00 equiv.) in CH2Cl2 (44.0 ml) was added dropwise to a solution of N,N-dimethylformamide (DMF) (18.9 ml, 244 mmol, 2.60 equiv.) in CH2Cl2 (90 ml) at room temperature under argon. After the reaction mixture was stirred at the same temperature for 1 h, it was cooled to −15 °C. A solution of 3,5-dimethoxyphenol (14.5 g, 93.8 mmol, 1.00 equiv.) in CH2Cl2 (58 ml) was added dropwise to the above reaction mixture for over 45 min. This mixture was stirred at −15 °C for 30 min and then at room temperature for 30 min. Next, 1 M aqueous HCl (130 ml) was added at 0 °C. The mixture was heated to 40 °C after stirring for 2 h at room temperature. After 3 h of stirring at the same temperature, the reaction mixture was added to a mixture of brine and EtOAc at 0 °C. The aqueous layer was extracted with two portions of EtOAc. The combined organic layers were washed with brine, dried over MgSO4 and concentrated in vacuo. The residue was re-crystallized from CH2Cl2-hexane to afford 16 as a white solid.7
Thin-layer chromatography, Rf=0.57 (hexane/ethyl acetate=1/1); 1H NMR (CDCl3) 12.5 (s, 1H), 10.1 (s, 1H), 6.00 (d, J=2.2 Hz, 1H), 5.91 (d, J=2.2Hz, 1H), 3.84 (s, 3H), 3.83 (s, 3H); 13C NMR (CDCl3) 191.9, 168.2, 166.4, 163.6, 106.1, 93.0, 90.6, 55.8; FT-IR (solid) 2984, 1624, 1498, 1475, 1211, 1157, 1111, 1045 cm−1.
4,6-Bis(benzyloxy)-2-hydroxybenzaldehyde (17)
K2CO3 (9.10 g, 65.8 mmol, 2.40 equiv.) was added to a solution of 2,4,6-trihydroxybenzaldehyde (5.30 g, 27.9 mmol, 1.00 equiv.) in DMF (70 ml) at room temperature under argon. The mixture was stirred at the same temperature for 5 min, after which benzyl bromide (6.64 g, 55.8 mmol, 2.00 equiv.) was added to it. After stirring for 5 h, the reaction mixture was added to a mixture of 1 M aqueous HCl and EtOAc at 0 °C. The aqueous layer was extracted with two portions of EtOAc. The combined organic layers were washed with three portions of H2O and brine, dried over MgSO4 and concentrated in vacuo. The residue was re-crystallized from CH2Cl2-hexane to afford 17 as a white solid.8
Results and discussion
Screening for inhibitors of glucose production in hepatic cells
We have developed a cell-based assay system for detecting hepatic glucose production by using the rat hepatoma cell line H4IIE-C3, which mimics in vivo liver function.9, 10 In this system, after culturing overnight in the serum-free medium, the cells were incubated in a glucose-free medium for 5 h and the amount of glucose accumulated in the medium was detected by glucose oxidase reaction. Glucose oxidase catalyzes the formation of D-gluconolactone and H2O2 from D-glucose; and H2O2 reacts with the Amplex Red reagent in the presence of horseradish peroxidase to generate a red fluorescent oxidation product, resorufin. Resorufin was quantified using a fluorescence microplate reader (Figure 1a). In this system, H4IIE-C3 cells produced glucose, which was accumulated in the medium at a concentration of 25 μM. When metformin was added into the glucose-free medium as a positive control for inhibition of hepatic glucose production, metformin inhibited the production of glucose in the medium with an IC50 value of 1.7 mM without cytotoxicity (IC50 >100 mM) (Figure 1b). This result suggests that this system is effective for screening inhibitors of glucose production.

Assay system and the screening of inhibitors of hepatic glucose production. (a) Flow chart of the assay system of glucose production in rat liver cells. (b) Inhibitory effect of metformin on glucose production. (c) Use of this system in screening inhibitors of glucose production from the natural product library.
In the screening process, samples to be screened were added in the glucose-free medium. The samples that showed ⩾55% inhibition of glucose production compared with the glucose production in the medium to which samples were not added were selected as inhibitors of glucose production. A total of 40 508 natural product samples were used for the primary screening. We screened all the samples at a final concentration of 1% in a volume of 20 μl per well using a 384-well format. The first screenings yielded 566 hit samples (1.40%). Among the 566 hit samples, some of them probably behaved as inhibitors of glucose oxidase and horseradish peroxidase in the glucose assay system, some acted as quenchers of resorufin, and some as scavengers of H2O2. Hence, we excluded samples that caused a decrease in the fluorescence intensity when 50 μM of glucose solution was separately added into the same assay system (fluorescence intensity, <70%). Furthermore, we excluded samples that showed cytotoxicity during 5 h of incubation (cytotoxicity, >40%). After performing additional reproducible and dose-dependent tests, we finally selected five samples (0.012%) as inhibitors of glucose production. Four of these were 35, 46, 55 and 6, which were selected from the library of isolated microbial metabolites. The remaining sample was obtained from the culture extract of Fusarium sp. no. 351386. Therefore, we cultured this fungus in fermentation media and isolated two active compounds—13 and 24—from its culture extract.
Identification of 1 and 2
The structures of the active compounds, 1 and 2, were determined by HR-ESI-MS, UV and 1H NMR spectra as follows: Javanicin, 1H NMR (CDCl3): δ 13.24. (s, 1H), 12.84 (s, 1H), 6.19 (s, 1H), 3.92 (s, 3H), 3.89 (s, 2H), 2.28 (s, 3H), 2.22 (s, 3H). UV λmax 536, 501, 472, 303; HR-MS m/z 291.0868 (calcd for C15H15O6, [M+H]+ 291.0869). Solaniol, 1H NMR δ (CDCl3): 13.31 (s, 1H), 13.04 (s, 1H), 6.19 (s, 1H), 4.11 (m, 1H), 3.91 (s, 3H), 2.93 (d, 2H, J=5.5 Hz), 2.33 (s, 3H), 1.30 (d, 3H, J=6.0 Hz). UV λmax 540, 504, 475, 307; HR-MS m/z 293.1052 (calcd for C15H17O6, [M+H]+ 293.1025).
Inhibition of hepatic glucose production by naphthoquinone derivatives
We tested the activities of five naphthoquinone derivatives with regard to inhibition of hepatic glucose production. The structures of 3–5 revealed that they were derivatives of 1 and 2, as shown in Table 1. All these compounds exhibited inhibitory effects on glucose production. Thus, the naphthoquinone moiety, common to all these compounds, may be essential for their inhibitory activities. The compounds 1–4 showed similar degrees of inhibition (IC50=3.8–6.8 μM); however, the inhibitory activity of 5 (IC50=30 μM) was approximately sixfold lower than the inhibitory activities of 1–4. Therefore, the pyridine moiety of 5 may have a significant role in inhibiting glucose production. In addition, these compounds, except 5, exhibited cytotoxic effects (IC50=3.3–8.8 μM) on H4IIE-C3 cells during 48 h of incubation, and 5 resulted in 30% inhibition of cell growth at a concentration of 50 μM. Hence, the IC50 values for the cytotoxic activity of these compounds were comparable with those of the inhibitory activity against hepatic glucose production (Table 1). On the basis of these results, it was concluded that these compounds are unsuitable candidates for use in anti-diabetic drugs.
Inhibition of hepatic glucose production by vanillin derivatives
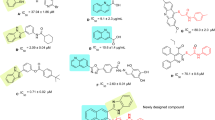
Additional screening showed that 6 from our library was also an inhibitor of glucose production. To examine structure–activity relationship, we compared the inhibitory effects of 6 derivatives 7–17 on glucose production (Table 2). All 6 derivatives inhibited hepatic glucose production and their IC50 values (IC50=24–190 μM) were lower than that of metformin (IC50=1.7 mM). In particular, 9 and 13 showed strong inhibitory activities. Moreover, these compounds did not exhibit cytotoxic effects on H4IIE-C3 cells at 50 μM after 48 h of incubation. Thus, 6 derivatives can be considered as suitable candidates for use in anti-diabetic drugs, as opposed to the naphthoquinone derivatives examined here.
The inhibitory activity of 7 (IC50=190 μM) against glucose production was weaker than that of 6 (IC50=32 μM), 8 (IC50=75 μM), 9 (IC50=25 μM) and 10 (IC50=39 μM) (Table 3). These results suggest that the absence of a carboxyl group in their structure was important for the inhibitory activity. On comparing the inhibitory activities of 5-halogenated salicylaldehydes (11, 12) and 5-nitrosalicylaldehyde (13), it was found that 13, which contains a 5-nitro group, showed the strongest inhibitory activity at an IC50 value of 24 μM. The activities of 14, 15, 16 and 17 were lower than those of 6, 11 and 13. Accordingly, in the case of benzaldehyde, 4-hydroxy-3-methoxy or 2,5-disubstituted moieties may have important roles in the inhibitory activities of these compounds against glucose production.
In this study, we developed a screening system for inhibitors of glucose production. Using the rat hepatoma cell line H4IIE-C3, we screened a total of 40 508 samples and evaluated 12 vanillin derivatives. We found that 9 or 13 inhibits glucose production in hepatic cells without exhibiting cytotoxic effects. In addition to these characteristics, these compounds also satisfy Lipinski's rule of 511 for oral drug-like properties as follows: a molecular weight of a compound is <500, hydrogen bond donors are <5, hydrogen bond accepters are <10 and an octanol–water partition co-efficient log P is <5. Therefore, 9 or 13 may have the potential to become leading oral anti-diabetic drugs.
Conflict of interest
The authors declare no conflict of interest.
References
Agius, L. New hepatic targets for glycaemic control in diabetes. Best Pract. Res. Clin. Endocrinol. Metab. 21, 587–605 (2007).
Scarpello, J. H. & Howlett, H. C. Metformin therapy and clinical uses. Diab. Vasc. Dis. Res. 5, 157–167 (2008).
Arnstein, H. R. V. & Cook, A. H. Production of antibiotics by fungi. Part III. Javanicin. An antibacterial pigment from Fusarium javanicum. J. Chem. Soc. 1021–1028 (1947).
Arsenault, G. P. Fungal metabolites- III Quinones from Fusarium solani D2 purple and structure of (+)-solaniol. Tetrahedron 24, 4745–4749 (1968).
Tatum, J. H., Baker, R. A. & Berry, R. E. Naphthoquinones produced by Fusarium oxysporum isolated from citrus. Phytochemistry 24, 457–459 (1985).
Tatum, J. H., Baker, R. A. & Berry, R. E. Metabolites of Fusarium solani. Phytochemistry 28, 283–284 (1989).
Roelens, F. et al. Regioselective synthesis and estrogenicity of (±)-8-alkyl-5,7-dihydroxy-4-(4-hydroxyphenyl)-3,4-dihydrocoumarins. Eur. J. Med. Chem. 40, 1042–1051 (2005).
Anderson, J. C., Headley, C., Stapleton, P. D. & Taylor, P. D. Asymmetric total synthesis of B-ring modified ()-epicatechin gallate analogues and their modulation of â-lactam resistance in Staphylococcus aureus. Tetrahedron 61, 7703–7711 (2005).
Hofmann, C., Marsh, J. W., Miller, B. & Steiner, D. F. Cultured hepatoma cells as a model system for studying insulin processing and biologic responsiveness. Diabetes 29, 865–874 (1980).
de Raemy-Schenk, A. M. et al. A cellular assay for measuring the modulation of glucose production in H4IIE cells. Assay Drug Dev. Technol. 4, 525–533 (2006).
Lipinski, C. A., Lombardo, F., Dominy, B. W. & Feeney, P. J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv Drug Deliv. Rev. 46, 3–26 (2001).
Acknowledgements
This work was supported by a grant from the New Energy and Industrial Technology Development Organization (NEDO), Japan.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Hashimoto, J., Motohashi, K., Sakamoto, K. et al. Screening and evaluation of new inhibitors of hepatic glucose production. J Antibiot 62, 625–629 (2009). https://doi.org/10.1038/ja.2009.93
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ja.2009.93
Keywords
This article is cited by
-
Two New Anthraquinones from the Marine-Derived Fungus Fusarium sp. J3-2
Chemistry of Natural Compounds (2023)
