
Abstract
The order and timing of species arrival during community assembly can have long term effects on community structure due to priority effects. The importance of such processes in complex bacterial communities where dispersal involves mixing of entire communities is currently not known. Here we used a transplant experiment with two bacterioplankton communities of different origin (freshwater and brackish). Sterile medium of each origin was initially inoculated with a bacterial community of different (‘alien’) origin, followed by dispersal of the respective ‘home’ community at different time points after initial inoculation. We found that the later the dispersal with the ‘home’ community occurred the smaller the effect on the final community composition. This suggests that priority effects by the initially inoculated community reduce the establishment success of taxa from the later arriving community and that this effect depends on dispersal timing.
Similar content being viewed by others

Main
The order and timing of species arrival during community assembly can have long term effects on community structure and functioning due to priority effects (Fukami et al., 2010; Dickie et al., 2012). Priority effects emerge when a species that arrives early exhausts or modifies available resources, and by that affects the establishment success of later arriving species (Shulman et al., 1983; Fukami, 2015). In addition, rapid adaptation of communities to new environmental conditions (Rainey and Travisano, 1998; Fukami et al., 2007) might further enforce priority effects (De Meester et al., 2016). Priority effects are potentially important wherever and whenever empty niche space becomes available (for example, during or after a disturbance; Fukami, 2015).
Experimental work has primarily been done with relatively simple model communities, and showed that priority effects are important for different organisms (Jiang and Patel, 2008; Altermatt et al., 2011; Hiscox et al., 2015). Although dispersal of single species that arrive in a location with clearly separated arrival sequences is possible, dispersal events in bacterial communities in nature are complex and involve mixing or coalescence of entire communities (Rillig et al., 2015). Since it is possible that single and multiple species invasion models give different results (Hewitt and Huxel, 2002), conclusions derived from work on simple model communities do not allow to make predictions about priority effects in complex natural communities. Therefore, we investigated the role of priority effects in bacterial communities of natural origin in a microcosm experiment in batch culture mode. We used water and their respective bacterial communities from 2 habitats that differed in environmental characteristics (lake water vs. brackish water from the Baltic Sea). Here the basic assumption is that communities from different origins are adapted to their respective environmental conditions. For both habitats, we prepared sterile medium as well as a bacterial inoculum and subsequently inoculated each medium with each inoculum (for full methods, see Supplementary Information). Thus, each medium was inoculated with an ‘alien’ community (that is, lake bacteria in brackish medium and vice versa) and inoculations with ‘home’ communities in their respective media (for example, lake bacteria in lake medium) served as controls. Following, we manipulated the timing of dispersal events with ‘home’ bacteria at constant rates 0, 6, 12, 24, 48, 72 and 96 h after the first inoculation, to all treatments (except 2 treatments which did not receive any dispersal—‘WD’ treatments). Community composition was determined by Illumina sequencing of the 16 S rRNA gene 1 and 2 weeks after the first inoculation (see Supplementary File for details). We hypothesize that (1) priority effects reduce the establishment success of taxa from the later arriving community. Hence, the stronger the priority effect, the greater the difference between cultures inoculated with ‘alien’ communities and their respective ‘home’ cultures (that is, controls); (2) the importance of priority effects depends on dispersal timing so that dissimilarities of ‘alien’ compared to the ‘home’ communities should be the greater the later the dispersal with the ‘home’ community occurs (Figure 1).

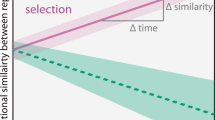
Conceptual figure showing three possible scenarios for the relationship between the dissimilarity of cultures inoculated with ‘alien’ compared to ‘home’ communities in dependence of the time of dispersal by the ‘home’ community: (a) a weak priority effect results in negligible differences between communities; (b) a strong priority effect dampens the establishment success of taxa from the later arriving community resulting in substantial difference between communities; (c) the importance of priority effects depends on dispersal timing so that dissimilarities between communities should be the greater the longer the time-lag between inoculation and dispersal with the ‘home’ community.
Regarding our first hypothesis, the ‘WD’ treatments in most cases had a greater dissimilarity compared to the dispersal treatments (Figure 2). This shows that dispersal by the potentially better adapted ‘home’ communities had the ability to influence community structure significantly. However, priority effects were nevertheless always strong enough to prevent complete convergence of the community mixes towards the ‘home’ communities (that is, Bray–Curtis dissimilarity>0, Figure 2). In general, priority effects were weaker (that is, smaller Bray–Curtis dissimilarities) in the brackish compared to lake medium (Wilcoxon paired test: P<0.0001) and after two compared to one week after inoculation (Wilcoxon paired test: P<0.0001; Figure 2). A possible explanation for the former could be that the brackish communities harbor a larger stock of freshwater bacteria as a result of the influx of freshwater bacteria via rivers and salt tolerant bacteria than vice versa. Accordingly, brackish communities have been shown to reach similar carrying capacities compared to the ‘home’ community (see Supplementary Information and Langenheder et al., 2003) and this higher versatility may therefore have resulted in overall greater dissimilarities in the freshwater medium.

Dissimilarities of cultures initially inoculated with ‘alien’ communities compared to those inoculated with their respective ‘home’ communities (that is, controls) for Baltic (a, c) and lake (b, d) medium 1 (a, b) and 2 (c, d) weeks after initial inoculation. Individual box-plots represent the timing of the dispersal events with ‘home’ bacteria (‘WD’—treatments without dispersal of ‘home’ community). Values of dissimilarity significantly different from ‘WD’ (Mann–Whitney test P<0.05) are indicated with asterisks. r and the P-values from linear correlations (computed without ‘WD’ treatments) are indicated in the graphs.
In support of our second hypothesis, we found that the dissimilarities of treatments inoculated with ‘alien’ communities compared to the respective controls were the greater the later the dispersal with the ‘home’ community occurred (effect of dispersal timing of ‘home’ communities: Permanova pseudo-F=7.215, P<0.0001; see Supplementary Table 3 for pair-wise comparisons; Figure 2). This suggests that priority effects were the stronger the later the dispersal with the better adapted ‘home’ community occurred (potentially as a result of competition for limited resources). Moreover, the effect of dispersal timing was stronger after 2 compared to 1 week (Figure 2). Hence, even though priority effects overall tended to decrease with time (see above), they remained stronger at the later time point in treatments where ‘alien’ communities were given more time to establish before the dispersal with the ‘home’ community occurred. However, longer experiments than 2 weeks are needed to differentiate whether the effect of priority effects on community structure are permanent or transient (Fukami, 2015). Moreover, also higher dispersal rates than those applied here (~20%) may in principle overrule the priority effects due to mass effects (De Meester et al., 2016), even though there is no evidence that this is the case (Pu and Jiang, 2015).
To conclude, previous studies have shown that high dispersal rates in combination with local environment influence the composition of natural bacterial communities (Lindström and Östman, 2011; Souffreau et al., 2014; Evans et al., 2016; Zha et al., 2016). Moreover, recent studies have pointed out the importance of an historical perspective on bacterial community assembly (Andersson et al., 2014; Vass and Langenheder, 2017). Our study shows for the first time that the timing of community coalescence events can have a pronounced influence on community structure due to priority effects, which stresses the importance of dispersal history for community assembly. Future studies including longer time-periods as well various environmental gradients and dispersal rates should now be done to ascertain the generality of priority effects in natural bacterial communities we observed here.
References
Altermatt F, Bieger A, Carrara F, Rinaldo A, Holyoak M . (2011). Effects of connectivity and recurrent local disturbances on community structure and population density in experimental metacommunities. PLoS One 6: e19525.
Andersson MGI, Berga M, Lindström ES, Langenheder S . (2014). The spatial structure of bacterial communities is influenced by historical environmental conditions. Ecology 95: 1134–1140.
De Meester L, Vanoverbeke J, Kilsdonk LJ, Urban MC . (2016). Evolving perspectives on monopolization and priority effects. Trends Ecol Evol 31: 136–146.
Dickie IA, Fukami T, Wilkie JP, Allen RB, Buchanan PK . (2012). Do assembly history effects attenuate from species to ecosystem properties? A field test with wood‐inhabiting fungi. Ecol Lett 15: 133–141.
Evans S, Martiny JBH, Allison SD . (2016). Effects of dispersal and selection on stochastic assembly in microbial communities. ISME J 11: 176–185.
Fukami T, Beaumont HJE, Zhang XX, Rainey PB . (2007). Immigration history controls diversification in experimental adaptive radiation. Nature 446: 436–439.
Fukami T, Dickie IA, Paula Wilkie J, Paulus BC, Park D, Roberts A et al. (2010). Assembly history dictates ecosystem functioning: evidence from wood decomposer communities. Ecol Lett 13: 675–684.
Fukami T . (2015). Historical contingency in community assembly: integrating niches, species pools, and priority effects. Annu Rev Ecol Evol Syst 46: 1–23.
Hewitt CL, Huxel GR . (2002). Invasion success and community resistance in single and multiple species invasion models: do the models support the conclusions? Biol Invasions 4: 263–271.
Hiscox J, Savoury M, Müller CT, Lindahl BD, Rogers HJ, Boddy L . (2015). Priority effects during fungal community establishment in beech wood. ISME J 9: 2246–2260.
Jiang L, Patel SN . (2008). Community assembly in the presence of disturbance: a microcosm experiment. Ecology 89: 1931–1940.
Langenheder S, Kisand V, Wikner J, Tranvik LJ . (2003). Salinity as a structuring factor for the composition and performance of bacterioplankton degrading riverine DOC. FEMS Microbiol Ecol 45: 189–202.
Lindström ES, Östman Ö . (2011). The importance of dispersal for bacterial community composition and functioning. PLoS One 6: e25883.
Pu Z, Jiang L . (2015). Dispersal among local communities does not reduce historical contingencies during metacommunity assembly. Oikos 124: 1327–1336.
Rainey PB, Travisano M . (1998). Adaptive radiation in a heterogeneous environment. Nature 394: 69–72.
Rillig MC, Antonovics J, Caruso T, Lehmann A, Powell JR, Veresoglou SD et al. (2015). Interchange of entire communities: microbial community coalescence. Trends Ecol Evol 30: 470–476.
Shulman MJ, Ogden JC, Ebersole JP, McFarland WN, Miller SL, Wolf NG . (1983). Priority effects in the recruitment of juvenile coral-reef fishes. Ecology 64: 1508–1513.
Souffreau C, Pecceu B, Denis C, Rummens K, De Meester L . (2014). An experimental analysis of species sorting and mass effects in freshwater bacterioplankton. Freshwater Biol 59: 2081–2095.
Vass M, Langenheder S . (2017). The legacy of the past: effects of historical processes on microbial metacommunities. Aquat Microb Ecol 79: 13–19.
Zha Y, Berga M, Comte J, Langenheder S . (2016). Effects of dispersal and initial diversity on the composition and functional performance of bacterial communities. PloS One 11: e0155239.
Acknowledgements
We thank the anonymous reviewers for their comments that led us to the improvement of the manuscript.
Author information
Authors and Affiliations
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on The ISME Journal website
Supplementary information
Rights and permissions
About this article

Cite this article
Svoboda, P., Lindström, E., Ahmed Osman, O. et al. Dispersal timing determines the importance of priority effects in bacterial communities. ISME J 12, 644–646 (2018). https://doi.org/10.1038/ismej.2017.180
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ismej.2017.180
This article is cited by
-
Routes and rates of bacterial dispersal impact surface soil microbiome composition and functioning
The ISME Journal (2022)
-
Farm-scale differentiation of active microbial colonizers
ISME Communications (2022)
-
Priority effects in microbiome assembly
Nature Reviews Microbiology (2022)
-
Repeated introduction of micropollutants enhances microbial succession despite stable degradation patterns
ISME Communications (2022)
-
Meter-scale variation within a single transect demands attention to taxon accumulation curves in riverine microbiome studies
Frontiers of Environmental Science & Engineering (2022)
