
Abstract
Background:
Obesity is an independent risk factor for morbidity and mortality from pandemic influenza H1N1. Influenza is a significant public health threat, killing an estimated 250 000–500 000 people worldwide each year. More than one in ten of the world's adult population is obese and more than two-thirds of the US adult population is overweight or obese. No studies have compared humoral or cellular immune responses to influenza vaccination in healthy weight, overweight and obese populations despite clear public health importance.
Objective:
The study employed a convenience sample to determine the antibody response to the 2009–2010 inactivated trivalent influenza vaccine (TIV) in healthy weight, overweight and obese participants at 1 and 12 months post vaccination. In addition, activation of CD8+ T cells and expression of interferon-γ and granzyme B were measured in influenza-stimulated peripheral blood mononuclear cell (PBMC) cultures.
Results:
Body mass index (BMI) correlated positively with higher initial fold increase in IgG antibodies detected by enzyme-linked immunosorbent assay to TIV, confirmed by HAI antibody in a subset study. However, 12 months post vaccination, higher BMI was associated with a greater decline in influenza antibody titers. PBMCs challenged ex vivo with vaccine strain virus, demonstrated that obese individuals had decreased CD8+ T-cell activation and decreased expression of functional proteins compared with healthy weight individuals.
Conclusion:
These results suggest obesity may impair the ability to mount a protective immune response to influenza virus.
Similar content being viewed by others

Introduction
Influenza causes some three to five million cases of severe illness and 250 000–500 000 deaths every year around the world, even in the absence of a major pandemic.1 Obesity is a growing health concern of epidemic proportions in many countries;2 more than one in ten of the world's adult population is obese2 and more than two-thirds of the US adult population is overweight or obese.3 In addition to co-morbidities such as cardiovascular disease and diabetes, obesity itself is an immunosuppressive condition.4, 5, 6 During the recent pandemic of influenza A/H1N1/2009 (pH1N1), obesity was recognized for the first time as an independent risk factor for increased influenza morbidity and mortality.7, 8, 9, 10, 11
Annual vaccination is the primary strategy available for decreasing the impact of influenza infection. No studies have examined how obesity may affect the response to influenza vaccination in humans. Obesity is associated with decreased antibody response to hepatitis B vaccine12, 13 and to tetanus toxoid.14 Our work with diet-induced obese mice found greater mortality following influenza infection and impaired innate immune responses,15 as well as, an impaired CD8+ T-cell memory response that increased morbidity and mortality from a secondary influenza challenge.16 However, the effects of obesity on immune responses to influenza vaccine have not been characterized in humans. We therefore initiated a prospective observational study of the effect of body mass index (BMI) on humoral and cell mediated immune responses to influenza vaccination in humans. Here, we report data from the first 2 years of this study.
Methods
Study design and subjects
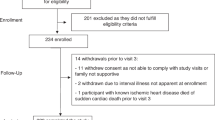
This is an ongoing, prospective observational study carried out at the University of North Carolina Family Medicine Center, an academic outpatient primary care facility in Chapel Hill, NC. Eligible participants were adult (⩾18 years) patients at the Center scheduled to receive the 2009–2010 seasonal trivalent influenza vaccine (TIV). Enrollment and data analysis were conducted independently for each year because of the annual change in vaccine composition. Exclusion criteria were immunosuppression, self-reported use of immunomodulator or immunosuppressive drugs, acute febrile illness, history of hypersensitivity to any influenza vaccine components, history of Guillian–Barre syndrome, or use of theophylline preparations or warfarin.17, 18 All procedures were approved by the Biomedical Institutional Review Board at the University of North Carolina.
In year 1 of the study (September–November 2009), we enrolled 499 participants. At enrollment, informed consent, height, weight and a baseline serum sample were obtained. One dose of 2009–2010 seasonal TIV (0.5 ml Fluzone (Sanofi Pasteur, Swiftwater, PA, USA) containing A/Brisbane/59/2007 (H1N1), A/Brisbane/10/2007 (H3N2) and B/Brisbane/60/2008) was administered in the deltoid muscle. Participants (461, 92% completion rate) returned 28–35 days later for a post-vaccination blood draw. Pre- and post-vaccination serum samples were stored at −80 °C until analyzed, whereas peripheral blood mononuclear cells (PBMCs) were separated from a second heparinized blood sample on a Histopaque (Sigma, St Louis, MO, USA) gradient and frozen in liquid nitrogen until analyzed.
At the start of year 2 of the study, 74 participants who had participated in year 1 independently re-enrolled to participate in year 2, providing us the additional opportunity to assess immune parameters 12 months after receipt of the 2009–2010 TIV.
ELISA and haemagglutination inhibition assays
IgG antibodies were quantified by enzyme-linked immunosorbent assay using the 2009–2010 seasonal TIV as antigen. Vaccine was diluted and adsorbed to microtitration plates in a carbonate coating buffer. After washing, triplicate serum dilutions in PBS were allowed to react with antigen, and bound antibodies were detected by a peroxidase-conjugated goat anti-human IgG (Abcam, Cambridge, MA, USA), followed by a chromogenic substrate. Color intensity was measured by absorbance at 450 nm. Internal control sera were included in each run. Pre- and post-vaccination sera from each participant were tested in the same run. The intra-assay coefficient of variation using this assay is 4%.
In order to determine serotype-specific antibody responses to the three components of the seasonal TIV, a subset of pre- and 1 month post-vaccine serum samples from 38 healthy weight and 38 obese participants, matched for race, age and sex, was tested by Focus Diagnostics (Cypress, CA, USA) for HAI antibodies against the 2009–2010 seasonal TIV strains.
Activation and expression of IFNγ and granzyme B by influenza-stimulated PBMCs
PBMC samples obtained from 61 participants 12 months post vaccination (23 healthy weight (BMI 22.2 +/− 1.7), 17 overweight (BMI 27.6 +/− 1.5) and 21 obese (BMI 35.7 +/− 4.5)) were thawed and cultured in AIM-V serum-free media supplemented with 1% penicillin/streptomycin and 1% glutamine. We have previously determined that our freezing method did not alter lymphocyte cell subset numbers in samples from healthy weight or obese individuals (data not shown). Cells were plated at 1 × 106 cells per well for 120 h with or without stimulation with 6.4 hemagglutination units of influenza A/Brisbane/59/2007 (H1N1). For the last 6 h, GolgiPlug, a protein transport inhibitor containing Brefeldin A, (BD Biosciences, San Jose, CA, USA) was added to all PBMC samples. PBMCs were stained with fluorochrome-conjugated antibodies and analyzed for CD3, CD8, CD69, interferon-γ (IFNγ) and granzyme B expression, using a Cyan ADP flow cytometer (Beckman Coulter, Fullerton, CA, USA). Appropriate isotype controls were used for each stain and data were analyzed using FlowJo software (TreeStar, Ashland, OR, USA). CD69 is an early-activation marker expressed on the surface of activated lymphocytes. CD8+ cytotoxic T cells function to destroy infected cells and mediate this targeted killing through a variety of mechanisms, including production of the functional proteins IFNγ and granzyme B.
Statistics
Associations between baseline variables and antibody response (at 1 month and 12 months post vaccination) were assessed using Spearman's rank correlation coefficient for continuous variables (age and BMI) and the Kruskal–Wallis test for categorical variables (diabetes, race, sex and smoking status). Baseline variables that were marginally associated with ELISA antibody fold increase at 1 month post vaccination were included in a multivariate linear regression model, with log10 antibody fold increase as the response variable. For the matched pairs HAI substudy, response frequencies (defined as ⩾4-fold increase or seroconversion) between healthy weight and obese participants were compared using McNemar's test. Comparisons of HAI titer fold increase between healthy weight and obese participants were assessed by applying the Wilcoxon signed rank test to the log of the fold increase ratio between healthy weight and obese participants within a pair. Fold increase between paired unstimulated and stimulated PBMC expression of CD69, granzyme B and IFNγ was assessed using the Wilcoxon signed rank test.
Results
Characteristics of the study population
The demographics of the study population are presented in Table 1. Our participants were 29.7% healthy weight, 33.4% overweight and 35.5% obese. The subset of 74 participants who were studied 12 months after receipt of the 2009–2010 vaccine were demographically similar to the overall study population (Table 1).
ELISA and HAI titers 1 month post vaccination
As others have reported,19 age was negatively correlated with antibody response as measured by ELISA (P<0.001). BMI was positively correlated with antibody fold increase (P<0.001). Antibody response was higher in females (P=0.001), consistent with previous studies of gender effect on vaccine response.20 There was a marginally significant association between race and antibody response (P=0.01), suggesting that African Americans had slightly higher responses than whites. There was no association of antibody response with smoking status or diabetes. In a multivariate model of log10 antibody fold increase including BMI, age, race and sex, both BMI and age remained significant predictors of fold increase (P=0.002 and P<0.001, respectively), whereas race was marginally significant (P=0.03) and sex was borderline significant (P=0.08). Based on the fitted model, a 10 unit increase in BMI was associated with 13% greater-fold increase in antibody titer, whereas a 10 year increase in age was associated with an 8% lower-fold increase in antibody titer. Age and BMI did not interact in the model.
Pre- and post-vaccination antibody titers for the three vaccine strains were not significantly different between the healthy weight and obese participants. As shown in Figure 1, when examining fold increase between pre- and post-vaccination antibody titers, there was no difference for the A/Brisbane/59/2007 (H1N1) and A/Brisbane/10/2007 (H3N2) strains (P=0.014 and P=0.09, respectively), however, there was a higher-fold increase in obese compared with healthy weight participants for B/Brisbane/60/2008 (P=0.04). Spearman's rank correlations between the ELISA and HAI fold increases demonstrated that the ELISA was positively correlated with HAI results for all three of the vaccine strains: a correlation of 0.57 (P<0.001) for A/Brisbane/59, 0.50 (P<0.001) for A/Brisbane/10 and 0.52 (P<0.001) for B/Brisbane/60.

Obese participants do not have an impaired initial response to influenza vaccination. Boxplots of the fold increase of the geometric mean titers of HAI response for each vaccine strain. Wilcoxon signed rank test of fold increase of healthy weight vs obese: (a) A/Brisbane/59/2007, P=0.14; (b) A/Brisbane 10/2007, P=0.09; (c) B/Brisbane/60/2008, P=0.04. Healthy weight n=40, obese n=40.
ELISA and HAI titers 12 months following vaccination
As shown in Figure 2a, increasing BMI was associated with a larger drop in antibody titer to 2009–2010 seasonal TIV as measured by ELISA 12 months after vaccination. To confirm this finding using HAI, we tested 17 matched pairs of healthy weight (BMI 22.6 +/− 1.9) and obese participants (BMI 35.4 +/−5.4) 1 and 12 months post vaccination for HAI antibodies against each vaccine strain of virus. Although the majority of both healthy weight and obese vaccinated participants had a decrease in antibody titer during this interval, a larger percentage of obese participants had a four-fold or greater drop in HAI titer at 12 months compared with healthy weight participants (Figure 2b). Thus, these results indicate that obese individuals have a steeper decline in vaccine antibody over time compared with healthy weight individuals.

Obesity results in a greater decline of influenza antibodies. (a) Correlation between BMI and percent antibody drop of ELISA titers. As the BMI increases, the drop in antibody at 12 months postvaccination is increased. Spearman's rank correlation: r=0.29 (P=0.01), n=74. (b) More obese individuals have a >4-fold drop in HAI titer at 12 months post vaccination compared with healthy weight individuals (McNemar's test P=0.16 for A/Brisbane/59, P=0.32 for A/Brisbane/10 and P=0.03 for B/Brisbane/60). Healthy weight n=17, obese n=17.
Decreased activation of influenza-specific CD8+ T cells in PBMCs obtained from obese individuals
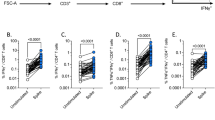
In order to test the cellular response to influenza vaccination, PBMCs at 12 months post vaccination were challenged ex vivo with live vaccine strain influenza A/Brisbane/59/2007 H1N1. PBMCs from obese participants exhibited a significantly lower percent increase in CD8+ T cells expressing the early activation marker CD69, than PBMCs from healthy weight participants (P=0.015) (Figure 3a), although the total numbers of CD8+ T cells were similar (data not shown).

Obesity results in defective CD8+ T-cell activation and production of the functional proteins Granzyme B and IFNγ by influenza-stimulated PBMCs. (a) PBMCs from obese participants have a lower-percent increase in activated CD69-expressing CD8+ T cells (P=0.015) and (b) a lower-pecentage increase in activated T cells that express Granzyme B (P=0.026) compared with healthy weight. (c) PBMCs from overweight and obese participants have a lower-percent increase in activated CD8+ T cells that express IFNγ (P=0.047 and P=0.006, respectively). The percent increase in cell number for each population of cells was calculated between PBMCs incubated with plain media and PBMCs incubated with influenza A virus. As such, each individual sample was compared with its own control. Bar graphs show mean percent increase and standard error for the three groups. Healthy weight n=23, overweight n=17, obese n=21. *indicates P-value is <0.05 compared with the healthy weight group. GrB=Granzyme B.
Decreased expression of functional proteins in influenza-specific activated CD8+ T cells in PBMCs obtained from obese individuals
In addition to upregulating activation markers upon stimulation, CD8+ T cells generate IFNγ and express granzyme B in order to limit influenza replication and rapidly clear the virus. PBMCs from obese participants exhibited a significantly lower-percent increase in activated CD8+ T cells expressing granzyme B than PBMCs from healthy weight participants (P=0.026) (Figure 3b). PBMCs from obese and overweight participants exhibited a lower-percent increase in activated CD8+ T cells expressing IFNγ, than PBMCs from healthy weight participants (P=0.006 and P=0.047, respectively) (Figure 3c). These data indicate that obesity and overweight in the case of IFNγ, results in a decreased production of the proteins IFNγ and granzyme B.
Discussion
During the 2009 H1N1 influenza pandemic, obesity was recognized as an independent risk factor for increased influenza morbidity and mortality.7, 8, 9 Influenza vaccination is the single most effective method for reducing morbidity and mortality from influenza. Despite recognition that obesity is immunosuppressive,4 this is the first study to examine antibody and CD8+ T-cell responses to influenza vaccination in healthy weight, overweight and obese individuals.
Because obesity reduces antibody responses to hepatitis B vaccine in adults and to tetanus vaccine in children,4, 12, 13, 14 elevated antibody response to influenza vaccination in our obese study participants was unexpected. Our data show that obese individuals mount a vigorous initial antibody response to TIV. However, a vaccine is protective only if the antibody titer is maintained throughout the period when influenza virus is circulating in the population. To examine the level of antibody maintenance after vaccination, we measured antibody levels 12 months after vaccination. Increases in BMI were positively correlated to decreases in antibody titer. More than 50% of the obese participants had a ⩾4-fold decrease in HAI titers to A/Brisbane/10 and B/Brisbane/60, and 47% had a ⩾4-fold decrease in HAI titer to A/Brisbane/59 at 12 months compared with 1 month post vaccination. By comparison, <25% of healthy weight participants had a >4-fold decrease in HAI titer to A/Brisbane/59 and B/Brisbane/60. The objectives of our ongoing study include more precise definition of the kinetics of this differential decline in antibody titer as well as follow-up of participants to determine whether BMI influences the actual rates of laboratory-confirmed influenza in vaccinated individuals. In addition to stimulating production of influenza antigen-specific antibodies, influenza vaccination also functions to generate a CD8+ T-cell response. The importance of a robust CD8+ T-cell memory response has been appreciated, and there is great interest in developing influenza vaccines that can promote a heightened T-cell memory response. Our own work in a murine diet-induced obesity model demonstrated an impaired CD8+ T-cell memory response leading to increased morbidity and mortality from an influenza challenge.16 In addition, it has also been suggested that there is an obesity-associated decrease in naive T cells and T-cell diversity,21 which could contribute to the impaired CD8+ T-cell response seen in our study.
Influenza-specific CD8+ T cells do not protect against infection, but instead act to limit progression of disease, allow for more rapid viral clearance and lessen the severity of disease.22 Whereas the targets for antibodies are the proteins on the surface of the influenza virus, the targets for CD8+ T cells are located on the internal, highly conserved proteins of the virus, which allow for extensive cross reactivity against multiple strains of influenza virus. The influenza virus surface proteins have a tendency to change frequently; as such, an antibody-based vaccine may be protective for 1 year, but not the next.23 Because memory CD8+ T cells are specific to internal influenza proteins that vary little from year to year, it is likely more effective and efficient to develop influenza vaccines which expand memory CD8+ T-cell populations, in addition to invoking a robust antibody response.24 Indeed, CD8+ T-cell cytotoxic activity correlates better with influenza protection than antibody titer in an elderly population.25
We found that percentage of influenza-activated CD8+ T cells were decreased in the obese participants, and two markers of functional CD8+ activity, IFNγ and granzyme B, were also significantly decreased in the obese participants. CD8+ T cells kill virus-infected cells by release of perforin and granzyme B26 and inhibit viral replication by the release of IFNγ.27 The fact that influenza-stimulated CD8+ T cells from obese individuals were deficient in the expression of both of these proteins strongly suggests that protection from an influenza infection may be not be optimal in the obese population.
We report here, for the first time, that influenza vaccine antibody levels decline significantly and CD8+ T-cell responses are defective in obese compared with healthy weight individuals. These findings suggest a mechanism for the increased risk of severe disease from pH1N1 infection in the obese population. If antibody titers and influenza vaccination-induced memory CD8+ T-cell populations are not maintained over time by the obese individuals, they may be at risk for suboptimal vaccine response. Additional studies are needed to determine the risk of influenza infection in a vaccinated obese population.
References
World Health Organization. WHO Factsheet 211: Influenza (Seasonal) [Internet]. April 2009 [cited 18 July 2011]. Available from: http://www.who.int/mediacentre/factsheets/fs211/en/index.html.
World Health Organization. WHO Factsheet 311: Obesity and Overweight [Internet]. March 2011 [cited 18 July 2011]. Available from: http://www.who.int/mediacentre/factsheets/fs311/en/index.html.
Catenacci VA, Hill JO, Wyatt HR . The obesity epidemic. Clin Chest Med 2009; 30: 415–444 vii.
Karlsson EA, Beck MA . The burden of obesity on infectious disease. Exp Biol Med (Maywood) 2010; 235: 1412–1424.
Huttunen R, Syrjanen J . Obesity and the outcome of infection. Lancet Infect Dis 2010; 10: 442–443.
Nave H, Beutel G, Kielstein JT . Obesity-related immunodeficiency in patients with pandemic influenza H1N1. Lancet Infect Dis 2011; 11: 14–15.
Center for Disease Control. Intensive-care patients with severe novel influenza A (H1N1) virus infection—Michigan, June 2009. Morb Mortal Wkly Rep 2009; 58: 749–752.
Louie JK, Acosta M, Samuel MC, Schechter R, Vugia DJ, Harriman K et al. A novel risk factor for a novel virus: obesity and 2009 pandemic influenza A (H1N1). Clin Infect Dis 2011; 52: 301–312.
Morgan OW, Bramley A, Fowlkes A, Freedman DS, Taylor TH, Gargiullo P et al. Morbid obesity as a risk factor for hospitalization and death due to 2009 pandemic influenza A(H1N1) disease. PLoS One 2010; 5: e9694.
Jain S, Kamimoto L, Bramley AM, Schmitz AM, Benoit SR, Louie J et al. Hospitalized patients with 2009 H1N1 influenza in the United States, April–June 2009. N Engl J Med 2009; 361: 1935–1944.
Van Kerkhove MD, Vandemaele KAH, Shinde V, Jaramillo-Gutierrez G, Koukounari A, Donnelly CA et al. Risk factors for severe outcomes following 2009 influenza A (H1N1) infection: a global pooled analysis. PLOS 2011; 8: e1001053.
Weber DJ, Rutala WA, Samsa GP, Santimaw JE, Lemon SM . Obesity as a predictor of poor antibody response to hepatitis B plasma vaccine. JAMA 1985; 254: 3187–3189.
Weber DJ, Rutala WA, Samsa GP, Bradshaw SE, Lemon SM . Impaired immunogenicity of hepatitis B vaccine in obese persons. N Engl J Med 1986; 314: 1393.
Eliakim A, Schwindt C, Zaldivar F, Casali P, Cooper DM . Reduced tetanus antibody titers in overweight children. Autoimmunity 2006; 39: 137–141.
Smith AG, Sheridan PA, Harp JB, Beck MA . Diet-induced obese mice have increased mortality and altered immune responses when infected with influenza virus. J Nutr 2007; 137: 1236–1243.
Karlsson EA, Sheridan PA, Beck MA . Diet-induced obesity impairs the T cell memory response to influenza virus infection. J Immunol 2010; 184: 3127–3133.
Patriarca P, Kendal AP, Stricof RL, Weber JA, Meissner MK, Dateno B . Influenza vaccination and warfarin or theophylline toxicity in nursing-home residents. N Engl J Med 1983; 308: 1601–1602.
Poli D, Chiarugi L, Capanni M, Antonucci E, Abbate R, Gensini GF et al. Need of more frequent international normalized ratio monitoring in elderly patients on long-term anticoagulant therapy after influenza vaccination. Blood Coagul Fibrinolysis 2002; 13: 297–300.
Goodwin K, Viboud, Simonsen L . Antibody response to influenza vaccination in the elderly: a quantitative review. Vaccine 2006; 24: 1159–1169.
Cook IF . Sexual dimorphism of humoral immunity with human vaccines. Vaccine 2008; 26: 3551–3555.
Yang H, Youm Y, Vandanmagsar B, Rood J, Kumar KG, Butler AA et al. Obesity accelerates thymic aging. Blood 2009; 114: 3803–3812.
Piet B, de Bree GJ, Smids-Dierdorp BS, van der Loos CM, Remmerswaal EB, von der Thüsen JH et al. CD8+ T cells with an intraepithelial phenotype upregulate cytotoxic function upon influenza infection in human lung. J Clin Invest 2011; 121: 2254–2263.
Carrat F, Flahault A . Influenza vaccine: the challenge of antigenic drift. Vaccine 2007; 25: 6852–6862.
Thomas PG, Keating R, Hulse-Post DJ, Doherty PC . Cell mediated protection in influenza infection. Emerg Infect Dis 2006; 12: 48–54.
Strassburg MA, Greenland S, Sorvillo FJ, Lieb LE, Habel LA . 1986 Influenza in the elderly: report of an outbreak and review of vaccine effectiveness reports. Vaccine 1986; 4: 38–44.
Russel JH, Ley TJ . Lymphocyte-mediated cytotoxicity. Ann Rev Immunol 2002; 20: 323–370.
Teijaro JR, Verhoeven D, Page CA, Turner D, Farber DL . Memory CD4T cells direct protective responses to influenza virus in the lungs through helper-independent mechanisms. J Virol 2010; 84: 9217–9226.
Acknowledgements
MAB received support from NIH RO1AI078090 and P30DK056350.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
This work is licensed under the Creative Commons Attribution-NonCommercial-No Derivative Works 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Sheridan, P., Paich, H., Handy, J. et al. Obesity is associated with impaired immune response to influenza vaccination in humans. Int J Obes 36, 1072–1077 (2012). https://doi.org/10.1038/ijo.2011.208
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ijo.2011.208
Keywords
This article is cited by
-
Antigen specific T cells in people with obesity at five months following ChAdOx1 COVID-19 vaccination
International Journal of Obesity (2023)
-
Accelerated waning of the humoral response to COVID-19 vaccines in obesity
Nature Medicine (2023)
-
Several areas of overlap between obesity and aging indicate obesity as a biomarker of accelerated aging of human B cell function and antibody responses
Immunity & Ageing (2022)
-
Immune and non-immune functions of adipose tissue leukocytes
Nature Reviews Immunology (2022)
-
The majority of SARS-CoV-2-specific antibodies in COVID-19 patients with obesity are autoimmune and not neutralizing
International Journal of Obesity (2022)
