
Abstract
Chymase is an ACE (angiotensin-converting enzyme)-independent angiotensin II-forming enzyme whose expression is increased in the maternal vascular endothelium in preeclampsia. However, mechanisms underlying chymase activation in preeclampsia remain unclear. Cathepsin C is a key enzyme in the activation of several serine proteases including chymase. In this study, we determined whether increased cathepsin C expression/activity might be responsible for the upregulation of chymase expression in preeclampsia. Maternal vascular cathepsin C, chymase and ACE expression were examined through immunohistochemical staining of subcutaneous fat tissue sections of normal and preeclamptic pregnant women. The role of cathepsin C in endothelial chymase and ACE expression was determined in cells treated with cathepsin C. Consequences of chymase activation were then determined by measurement of angiotensin II production in cells treated with the ACE inhibitor captopril and the chymase inhibitor chymostatin, separately and in combination. Expression of both cathepsin C and chymase, but not ACE expression, was markedly increased in the maternal vascular endothelium in subjects with preeclampsia compared with normal pregnant controls. Exogenous cathepsin C induced a dose-dependent increase in expression of mature cathepsin C and chymase, but not ACE, in endothelial cells. Moreover, angiotensin II production was significantly inhibited in cells treated with captopril or chymostatin alone and was further inhibited in cells treated with both inhibitors. These results suggest that cathepsin C upregulation induces chymase activation and subsequently promotes angiotensin II generation in endothelial cells. These data also provide evidence of upregulation of the cathepsin C–chymase–angiotensin signaling axis in maternal vasculature in preeclampsia.
Similar content being viewed by others

Introduction
Chymase is a chymotrypsin-like serine protease originally identified in mast cells.1 Chymase has a broad peptidolytic activity and is involved in a variety of cellular functions including cleavage of stem cell factor,2 stimulation of inflammatory cytokine production and induction of endothelial adhesion molecules.3, 4, 5 Chymase has important roles in the regulation of vascular function, through its ability not only to convert angiotensin I to angiotensin II, but also to convert big endothelin to endothelin-1.6, 7 It has been reported that chymase is responsible for ~70–80% of the angiotensin II generated in human heart tissue,6 and chymase is also a major producer of angiotensin-converting enzyme (ACE)-independent angiotensin II in human arteries.8 Consequently, it is widely accepted that chymase is a major non-ACE angiotensin II-generating enzyme in humans. Although the physiological role of chymase is still not understood, chymase has been implicated in the pathogenesis of several cardiovascular diseases, including heart failure, cardiac hypertrophy and diabetes mellitus.9 It has been speculated that activation of chymase may have profound effects on the vascular system because of its downstream vasoconstrictor-generating actions via angiotensin II and endothelin.10
Altered renin–angiotensin system activity is responsible for increased vascular resistance and has a significant role in the pathogenesis and pathophysiology of preeclampsia, a hypertensive disorder unique to human pregnancy. This notion is supported by several important findings. First, increased maternal vascular sensitivity to angiotensin II contributes to the development of preeclampsia.11, 12 Second, increased maternal levels of auto-antibody to angiotensin II receptor-1 are highly prevalent in women with preeclampsia.13 Third, expression and activity of chymase are increased not only in placental trophoblasts but also in the maternal systemic vasculature in preeclampsia.14, 15 Therefore, the activation of chymase is thought to significantly affect the renin–angiotensin system alterations associated with the increased angiotensin II generation and increased vasocontractility in preeclampsia. However, the mechanism by which chymase is activated in the vascular endothelium and placental trophoblasts in preeclampsia is largely unknown.
Cathepsin C, also known as dipeptidyl peptidase I (DPP-I), is a lysosomal exocysteine protease belonging to the peptidase C1 family. Cathepsin C is a key enzyme in the activation of many serine proteases in inflammatory cells.16, 17, 18, 19 Cathepsin C is essential for the activation of mast cell chymase in mice.18 In the present study, we investigated whether cathepsin C expression was upregulated in the maternal vasculature in women with pregnancy complicated by preeclampsia and determined whether increased cathepsin C expression might promote chymase activation and angiotensin II generation by vascular endothelial cells.
Materials and methods
Chemicals and reagents
Anti-human antibodies specific for cathepsin C (sc-74590) and ACE (sc-12187) were purchased from Santa Cruz (San Diego, CA, USA). An anti-chymase antibody (MA5-11717) was purchased from Thermo Scientific (Rockford, IL, USA). A β-actin antibody and angiotensin I were obtained from Sigma Chemicals (St Louis, MO, USA). Captopril and chymostatin were obtained from EMD Millipore (Darmstadt, Germany). All the other chemicals and reagents were obtained from Sigma Chemicals unless otherwise noted.
Sample collection
Maternal subcutaneous fat tissue was collected during cesarean section delivery from 12 pregnant women (six from normal and six from preeclampsia) and was used for examination of cathepsin C, chymase and ACE expression. The protocol for collection of subcutaneous fat tissue was approved by the IRB at Louisiana State University Health Sciences Center—Shreveport (LSUHSC-S), LA, USA. Normal pregnancy was defined as pregnancy with blood pressure <140/90 mm Hg, absence of proteinuria and the absence of obstetrical and medical complications. Diagnosis of preeclampsia was defined as follows: sustained systolic blood pressure of ⩾140 mm Hg or a sustained diastolic blood pressure of ⩾90 mm Hg on two separate readings; proteinuria measurement of 1+ or more on a dipstick, or 24 h urine protein collection with ⩾300 mg in the specimen. Smokers were excluded from the study, and no study subjects had signs of infection. To avoid clinical phenotypic differences in preeclamptic patients, patients with HELLP syndrome (hemolysis, elevated liver enzyme and low platelet count), diabetes and/or renal disease were excluded. None of the subjects in the preeclampsia group had a history of chronic hypertension. The clinical characteristics of the study subjects are presented in Table 1.
Immunohistochemistry
Freshly obtained subcutaneous fat tissue was immediately fixed in 10% formalin and then embedded in paraffin. Expression of cathepsin C, chymase and ACE was examined through immunohistochemistry in paraffin-embedded tissue sections. Stained slides were counterstained with Gill’s formulation hematoxylin. Tissue sections stained with secondary antibody only were used as controls. All slides stained with the same antibody were processed at the same time. Stained tissue slides were reviewed under an Olympus microscope, and images were captured.
Endothelial cell isolation and culture
Human umbilical cord vein endothelial cells (HUVECs) from normal-term placentas were isolated as previously described.20, 21 Isolated endothelial cells were cultured with endothelial growth medium (Lonza, Allendale, NJ, USA) supplemented with hydrocortisone, ascorbic acid, bovine brain extract, epidermal growth factor, gentamicin sulfate amphotericin (GA-1000) and fetal bovine serum. HUVECs at passage 1–2 were used for the experiments. Uterine myometrium vascular endothelial cells (UtMVECs) purchased from Lonza were also used in the study. UtMVECs were also incubated with endothelial growth medium (Lonza).
Protein expression
Protein expression of cathepsin C, chymase and ACE was determined by western blot analysis. Briefly, 10 μg of total cellular protein was subjected to electrophoresis (Bio-Rad, Hercules, CA, USA) and then transferred to nitrocellulose membranes. After being blocked, the membrane was probed with primary antibody (same antibody as used for IHC), followed by a relevant secondary antibody. An enhanced chemiluminescent detection kit (Amersham Corporation, Arlington Heights, IL, USA) and X-ray film were used to visualize the bound antibody. β-actin expression was determined for each sample. After scanning, the density of bands was analyzed with the NIH ImageJ analysis program. β-actin expression was used to normalize cathepsin C, chymase and ACE expression. Data are presented as the mean±s.e. from three independent experiments.
Angiotensin II measurement
Endothelial angiotensin II production was measured through an enzyme-linked immunoassay. The angiotensin II enzyme-linked immunoassay kit was purchased from Cayman Chemical (Ann Arbor, MI, USA) and consisted of angiotensin II standard, anti-angiotensin II IgG tracer, glutaraldehyde, borane trimethylamine, Ellman’s reagent, assay and wash buffers. The assay was performed according to the manufacturer’s instructions. The range of the standard curve was 0.98–125 pg ml−1. Medium samples were diluted 1:10 and measured in duplicate. Within-assay variation was <6%. The enzyme-linked immunoassay plate was read at 405 nm with an auto-plate reader (Molecular Devices, Sunnyvale, CA, USA).
Data analysis
Clinical demographic data are presented as the mean±s.d. Data for protein expression and angiotensin II production are presented as the mean±s.e. Statistical analysis was performed with Mann–Whitney test or analysis of variance with the computer software Prism 5 (GraphPad Software, La Jolla, CA, USA). The Student–Newman–Keuls test was used as a post hoc test. A probability level less than 0.05 was considered statistically significant.
Results
Cathepsin C and chymase expression are upregulated in the maternal vascular endothelium in preeclampsia
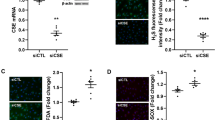
To determine whether altered cathepsin C expression is present in the maternal vascular endothelium in women with preeclampsia, cathepsin C expression was examined through immunohistochemical staining in tissue sections of subcutaneous fat tissues from normal and preeclamptic pregnant women. Our results showed that cathepsin C was expressed at only low levels in the endothelium of maternal vessels from normal pregnant women, but was strongly expressed in both the endothelium and the smooth muscle layer of vessels from women with preeclampsia (Figure 1). Because cathepsin C is a key protease that promotes chymase activation, and both chymase and ACE convert angiotensin I to angiotensin II, we also examined chymase and ACE expression in tissue sections from normal and preeclamptic pregnant women. The results were consistent with our previously reported finding that chymase is strongly expressed in vascular endothelium in samples from preeclamptic pregnant women compared with normal pregnant controls (Figure 1). ACE was expressed in both endothelial and vascular smooth muscle layers in vascular specimens but showed no difference between vessels from normal and preeclamptic pregnant women (Figure 1).

Representative images showing cathepsin C, chymase and ACE expression in maternal vessels of normal and preeclamptic pregnancies. (a, c, e) Normal pregnancies; (b, d, f) preeclampsia; (a, b) cathepsin C; (c, d) chymase; and (e, f) ACE. Scale bar=50micron. Increased cathepsin C and chymase expression was observed in maternal vascular endothelium from preeclampsia compared with normal pregnancy. Endothelial ACE expression was not different between normal and preeclamptic pregnancies.
Cathepsin C promotes expression of chymase but not ACE in endothelial cells
Because cathepsin C has a key role in inducing activation of many serine proteases, we next determined whether cathepsin C might promote chymase activation. In this experiment, confluent endothelial cells (HUVECs) grown in six-well plates were treated with different concentrations of cathepsin C for 2 h, and total cellular protein was collected. Protein expression for cathepsin C, chymase and ACE was determined by western blotting. The results are shown in Figures 2a and b. Figure 2a shows representative blots of cathepsin C, chymase and ACE expression. Two bands were detected for cathepsin C: a band at 55 kDa, which is the cathepsin C propeptide, and a band at 25 kDa, which is mature cathepsin C. Mature cathepsin C expression was markedly increased in cells treated with cathepsin C, and this cathepsin C-induced mature cathepsin C expression was increased in a dose-dependent manner, Figure 2b. Interestingly, chymase expression was also increased in a dose-dependent manner in cells treated with cathepsin C. However, ACE expression was not altered. The bar graphs show mature cathepsin C, chymase and ACE expression after normalization to β-actin expression for each sample (Figure 2b). Data are presented as the mean±s.e. from three independent experiments. The ratio of chymase to ACE expression was also calculated and it showed a dose-dependent increase in cells treated with cathepsin C in Figure 2b, thus indicating that cathepsin C modulates the relationship between chymase and ACE expression in endothelial cells.

Effects of cathepsin C on chymase and ACE expression in endothelial cells. Confluent endothelial cells were treated with different concentrations of cathepsin C. Total cellular protein was collected and protein expression for cathepsin C, chymase and ACE was determined by western blotting. (a) Protein expression of cathepsin C, chymase and ACE in endothelial cells treated with cathepsin C at concentrations of 0, 1, 5, 20 and 100 pg ml−1. (b) Bar graphs showing the mean±s.e. of cathepsin C, chymase and ACE expression from three independent experiments. The ratio of chymase to ACE expression was also calculated, *P<0.05 and **P<0.01: treated vs. untreated control. Exogenous cathepsin C induced expression of mature cathepsin C and chymase. Exogenous cathepsin C had no effect on ACE expression in endothelial cells. A full color version of this figure is available at the Hypertension Research journal online.
Cathepsin C mediated chymase activation and angiotensin II generation in endothelial cells
To further investigate whether cathepsin C-mediated chymase activation is responsible for increased angiotensin II generation by endothelial cells, we evaluated angiotensin II production in cells treated with cathepsin C in the presence of the ACE inhibitor captopril or the chymase inhibitor chymostatin. Two types of endothelial cells were used: HUVECs and UtMVECs. Our results showed that in HUVECs, at least 50% of the angiotensin II generated was inhibited by either captopril or chymostatin. Angiotensin II production was almost completely inhibited when both inhibitors were added to these cultures. In UtMVECs, a similar effect was found for captopril, but chymostatin alone inhibited ~80% of the angiotensin II produced (Figure 3, lower panel). These data suggested that chymase is a more potent angiotensin II-generating enzyme than ACE in UtMVECs.

Cathepsin C-mediated angiotensin II production in endothelial cells with or without pretreatment with the ACE inhibitor captopril and the chymase inhibitor chymostatin. Confluent endothelial cells were treated with 1 μM captopril or 100 μg ml−1 chymostatin for 2 h and then 100 pg ml−1 cathepsin C for 2 h followed by 10−7M angiotensin I for 30 min. The culture medium was then collected and analyzed for angiotensin II. Angiotensin II production was significantly decreased in cells pretreated with either captopril or chymostatin. Angiotensin II production was completely inhibited when both inhibitors were used in combination. The inhibitory effect was found in both HUVECs (upper panel) and UtMVECs (lower panel). **P<0.001: cathepsin C+Ang I vs. control and CC alone; ##P<0.01, captopril+cathepsin C+angiotensin I or chymostatin+cathepsin C+angiotensin I or captopril+chymostatin+cathepsin C+angiotensin I vs. cathepsin C+angiotensin I; ^P<0.05 and ^^P<0.01: captopril+chymostatin+cathepsin C+angiotensin I vs. captopril+Cathepsin C+angiotensin I or chymostatin+cathepsin C+angiotensin I in HUVECs and captopril+chymostatin+cathepsin C+angiotensin I vs. captopril+Cathepsin C+angiotensin I, but not chymostatin+cathepsin C+angiotensin I in UtMVECs. HUVEC, human umbilical cord vein endothelial cell; UtMVEC, uterine myometrium vascular endothelial cell.
Discussion
In the present study, we investigated potential mechanisms underlying chymase activation in endothelial cells and determined whether altered cathepsin C expression was present in the maternal vasculature in women with preeclampsia. Cathepsin C expression in maternal vascular endothelium was examined through immunohistochemical staining of subcutaneous fat tissue sections. Cathepsin C was scarcely detected in the maternal vessels of normal pregnant women. However, strong cathepsin C expression was detected not only in the vascular endothelium but also in the smooth muscle layer of vessels from women with preeclampsia. These findings clearly showed that cathepsin C expression is upregulated in the maternal vasculature of women with preeclampsia.
Cathepsin C is a key enzyme in the activation of many granule-associated serine proteases or peptidases in inflammatory cells, such as elastase and cathepsin G in neutrophils and chymase and tryptase in mast cells. We have previously reported that chymase expression is upregulated in the maternal vascular endothelium in women with preeclampsia.15 Because cathepsin C is a chymase activator, and chymase has a similar function as ACE in converting angiotensin I to angiotensin II, we also examined chymase and ACE expression in tissue sections from normal and preeclamptic pregnant women. We found that endothelial expression of chymase, but not ACE, was markedly increased in maternal vessels from preeclamptic women compared with normal pregnant women. Although the sample size is small in the present study, we have previously used a similar sample size to demonstrate that downregulation of expression of the anti-inflammatory mediator suppressor of cytokine signaling-3 is related to the upregulation of inflammatory microRNA-203 expression in the maternal vessel endothelium in preeclampsia.22, 23 A similar sample size has also been used by Shukla et al.24 to examine neutrophil infiltration and myeloperoxidase expression in subcutaneous fat vessels from normal and obesity pregnant women and by Mousa et al.25 to examine DNA methylation in maternal vessels of omental fat tissues from women with normal and preeclamptic pregnancies. Therefore, we believe that our finding of increased endothelial cathepsin C and chymase expression in women with preeclampsia is valid. Although information regarding maternal levels or activity of cathepsin C in normal pregnancy and preeclampsia is lacking, increases in the expression of both cathepsin C and chymase in the maternal vascular endothelium in preeclampsia suggest that cathepsin C upregulation may lead to chymase activation, which might subsequently trigger the vascular dysfunction seen in women with preeclampsia.
To determine whether cathepsin C might promote chymase expression in endothelial cells, we measured expression of cathepsin C, chymase and ACE in endothelial cells treated with different doses of cathepsin C. Interestingly, exogenous cathepsin C promoted mature cathepsin C production/expression and concurrent chymase upregulation in endothelial cells. Expression of mature cathepsin C and chymase was increased in a dose-dependent manner, whereas ACE expression was not affected. These data provided evidence of the intimate association between cathepsin C and chymase showing that cathepsin C activates chymase in endothelial cells. This concept is very important in terms of vascular regulation, especially angiotensin II-mediated vasocontractility, because chymase is a potent non-ACE angiotensin II-generating enzyme, and angiotensin II is a potent vasoconstrictor in humans. In addition, we observed that ACE expression was a dominant regulator in non-stimulated endothelial cells. This notion was supported by the observations of dominant expression of ACE in the maternal vascular endothelium in normal pregnant women and in cultured endothelial cells (in the absence of cathepsin C stimulation). However, this relationship between ACE and chymase expression was apparently reversed in maternal vessels during preeclampsia and in cells treated with cathepsin C, in which chymase expression dominated in maternal vessel endothelium in preeclampsia and an increased ratio of chymase to ACE expression was found in cells treated with cathepsin C. These observations led us to speculate that chymase might be the major angiotensin II-forming enzyme under conditions in which cathepsin C and chymase are activated.
To further characterize the consequences of cathepsin C and chymase activation in endothelial cells, we evaluated angiotensin II production in endothelial cells treated with cathepsin C in the presence of the ACE inhibitor captopril and the chymase inhibitor chymostatin. Our results showed that both captopril and chymostatin significantly inhibited angiotensin II generation in the presence of cathepsin C. Furthermore, this inhibitory effect on angiotensin II generation was further potentiated when both inhibitors, captopril and chymostatin, were used in combination rather than when either was used alone. These results suggested that vascular cathepsin C and chymase activation may be at least partly responsible for increased vasoconstriction through activation of the chymase–angiotensin II regulatory pathways in hypertensive disorders including preeclampsia.
Cathepsins are a family of proteases involved in the turnover of proteins, including extracellular matrix degradation. Cathepsins are distinguished by their structure, catalytic mechanism and the proteins that they cleave. Cathepsins A and G are serine proteases, cathepsins B, C, F and S are cysteine proteases and cathepsins D and E are aspartyl proteases. Cathepsins are also considered to be ‘inflammatory proteases’, and the increased activity of several of these proteases has been found in many inflammatory diseases, such as rheumatoid arthritis, chronic obstructive pulmonary disease, inflammatory bowel disease and asthma.26, 27, 28, 29, 30 Nakajima et al.31 have reported that maternal serum cathepsin D activity was higher in women with preeclampsia than in normal pregnant controls. Although little is currently known about altered cathepsin C levels in pregnancy and preeclampsia, activation of cathepsin C has been found to have a role in cardiovascular-related disorders. For example, Herías et al.32 have reported upregulation of cathepsin C gene and protein expression in atherosclerotic lesions in both men and mice. These investigators have further found that increased cathepsin C expression appears to be confined to inflammatory foci within the atherosclerotic plaque and is particularly associated within intimal foam cells and macrophages. In contrast, cathepsin C deficiency in monocytes and macrophages has been found to decrease plaque size in aortic atherosclerotic lesions.32 These findings suggest important contributions of this cysteine protease in the development of atherosclerosis. An animal study conducted by Pagano et al.33 has also shown that cathepsin C, through regulation of the production of the CXC-chemokine ligand, increases neutrophil recruitment during the development of experimental abdominal aortic aneurysms in mice. Moreover, increased cathepsin C activity has also been demonstrated in aortic artery tissues in renal-clip-induced hypertension in rats.34 Our findings of increased chymase expression associated with cathepsin C upregulation in the maternal vasculature in women with preeclampsia, with existing evidence of cathepsin C’s involvement in cardiovascular diseases in animal studies, indicate that dysregulation of this cysteine protease potently contributes to the development of cardiovascular disorders.
In conclusion, this is the first study to show that cathepsin C expression is upregulated in maternal vascular endothelium in women with preeclampsia. We further demonstrated that cathepsin C induces endothelial chymase activation, thus resulting in a discordant relationship between ACE and chymase in endothelial cells. Although the reason for the lack of stimulation of ACE expression in response to chymase is not known, this phenomenon is very interesting and may have a significant impact on the cardiovascular system, not only in preeclampsia but also in cardiovascular diseases in general, because both ACE and chymase are angiotensin II-generating enzymes. Therefore, it is reasonable to speculate that upregulation of the cathepsin C–chymase signaling pathway may have a critical role in increasing vasoconstriction and vascular dysfunction in preeclampsia. Moreover, the inhibitory effect of chymostatin against endothelial angiotensin II generation further suggests that this novel cathepsin C–chymase signaling axis may be a potential therapeutic target for the management of hypertension-related cardiovascular diseases.
References
Caughey GH, Zerweck EH, Vanderslice P . Structure, chromosomal assignment, and deduced amino acid sequence of a human gene for mast cell chymase. J Biol Chem 1991; 266: 12956–12963.
Longley BJ, Tyrrell L, Ma Y, Williams DA, Halaban R, Langley K, Lu HS, Schechter NM . Chymase cleavage of stem cell factor yields a bioactive, soluble product. Proc Natl Acad Sci USA 1997; 94: 9017–9021.
Mizutani H, Schechter N, Lazarus G, Black RA, Kupper TS . Rapid and specific conversion of precursor interleukin 1 beta (IL-1 beta) to an active IL-1 species by human mast cell chymase. J Exp Med 1991; 174: 821–825.
Wang Y, Zhang Y, Lewis DF, Gu Y, Li H, Granger DN, Alexander JS . Protease chymotrypsin mediates the endothelial expression of P- and E-selectin, but not ICAM and VCAM, induced by placental trophoblasts from preeclamptic pregnancies. Placenta 2003; 24: 851–861.
Gu Y, Liu C, Alexander JS, Groome LJ, Wang Y . Chymotrypsin-like protease (chymase) mediates endothelial activation by factors derived from preeclamptic placentas. Reprod Sci 2009; 16: 905–913.
Urata H, Kinoshita A, Misono KS, Bumpus FM, Husain A . Identification of a highly specific chymase as the major angiotensin II-forming enzyme in the human heart. J Biol Chem 1990; 265: 22348–22357.
D'Orléans-Juste P, Houde M, Rae GA, Bkaily G, Carrier E, Simard E . Endothelin-1 (1-31): from chymase-dependent synthesis to cardiovascular pathologies. Vascul Pharmacol 2008; 49: 51–62.
Richard V, Hurel-Merle S, Scalbert E, Ferry G, Lallemand F, Bessou JP, Thuillez C . Functional evidence for a role of vascular chymase in the production of angiotensin II in isolated human arteries. Circulation 2001; 104: 750–752.
Wu Q, Kou HC, Deng GG . Serine proteases and cardiac function. Biochim Biophys Acta 2005; 1751: 82–94.
Nakano A, Kishi F, Minami K, Wakabayashi H, Nakaya Y, Kido H . Selective conversion of big endothelins to tracheal smooth muscle-constricting 31-amino acid-length endothelins by chymase from human mast cells. J Immunol 1997; 159: 1987–1992.
Gant NF, Daley GL, Chand S, Whalley PJ, MacDonald PC . A study of angiotensin II pressor response throughout primigravid pregnancy. J Clin Invest 1973; 52: 2682–2689.
Gant NF, Chand S, Worley RJ, Whalley PJ, Crosby UD, MacDonald PC . A clinical test useful for predicting the development of acute hypertension in pregnancy. Am J Obstet Gynecol 1974; 120: 1–7.
Siddiqui AH, Irani RA, Blackwell SC, Ramin SM, Kellems RE, Xia Y . Angiotensin receptor agonistic autoantibody is highly prevalent in preeclampsia: correlation with disease severity. Hypertension 2010; 55: 386–393.
Wang Y, Gu Y, Zhang Y, Lewis DF, Alexander JS, Granger DN . Increased chymotrypsin-like protease (chymase) expression and activity in placentas from women with preeclampsia. Placenta 2007; 28: 263–269.
Wang Y, Gu Y, Lewis DF, Alexander JS, Granger DN . Elevated plasma chymotrypsin-like protease (chymase) activity in women with preeclampsia. Hypertens Pregn 2010; 29: 253–261.
Henningsson F, Wolters P, Chapman HA, Caughey GH, Pejler G . Mast cell cathepsins C and S control levels of carboxypeptidase A and the chymase, mouse mast cell protease 5. Biol Chem 2003; 384: 1527–1531.
Méthot N, Guay D, Rubin J, Ethier D, Ortega K, Wong S, Normandin D, Beaulieu C, Reddy TJ, Riendeau D, Percival MD . In vivo inhibition of serine protease processing requires a high fractional inhibition of cathepsin C. Mol Pharmacol 2008; 73: 1857–1865.
Wolters PJ, Pham CT, Muilenburg DJ, Ley TJ, Caughey GH . Dipeptidyl peptidase I is essential for activation of mast cell chymases, but not tryptases, in mice. J Biol Chem 2001; 276: 18551–18556.
Turk D, Janjić V, Stern I, Podobnik M, Lamba D, Dahl SW, Lauritzen C, Pedersen J, Turk V, Turk B . Structure of human dipeptidyl peptidase I (cathepsin C): exclusion domain added to an endopeptidase framework creates the machine for activation of granular serine proteases. EMBO J 2001; 20: 6570–6582.
Wang Y, Adair CD, Coe L, Weeks JW, Lewis DF, Alexander JS . Activation of endothelial cells in preeclampsia: increased neutrophil-endothelial adhesion correlates with up-regulation of adhesion molecule P-selectin in human umbilical vein endothelial cells isolated from preeclampsia. J Soc Gynecol Investig 1998; 5: 237–243.
Jaffe EA, Nachman RL, Becker CG, Minick CR . Culture of human endothelial cells derived from umbilical veins. Identification by morphologic and immunologic criteria. J Clin Invest 1973; 52: 2745–2756.
Wang Y, Lewis DF, Gu Y, Zhao S, Groome LJ . Elevated maternal soluble gp130 and IL-6 levels and reduced gp130 and SOCS-3 expressions in women with preeclampsia. Hypertension 2011; 57: 336–342.
Wang Y, Dong Q, Gu Y, Groome LJ . Up-regulation of miR-203 expression induces endothelial inflammatory response: potential role in preeclampsia. Am J Reprod Immunol 2016; 76: 482–490.
Shukla J, Walsh SW . Neutrophil release of myeloperoxidase in systemic vasculature of obese women may put them at risk for preeclampsia. Reprod Sci 2015; 22: 300–307.
Mousa AA, Archer KJ, Cappello R, Estrada-Gutierrez G, Isaacs CR, Strauss JF 3rd, Walsh SW . DNA methylation is altered in maternal blood vessels of women with preeclampsia. Reprod Sci 2012; 19: 1332–1342.
Adkison AM, Raptis SZ, Kelley DG, Pham CT . Dipeptidyl peptidase I activates neutrophil-derived serine proteases and regulates the development of acute experimental arthritis. J Clin Invest 2002; 109: 363–371.
Wolters PJ, Laig-Webster M, Caughey GH . Dipeptidyl peptidase I cleaves matrix-associated proteins and is expressed mainly by mast cells in normal dog airways. Am J Respir Cell Mol Biol 2000; 22: 183–190.
Hausmann M, Obermeier F, Schreiter K, Spottl T, Falk W, Schölmerich J, Herfarth H, Saftig P, Rogler G . Cathepsin D is up-regulated in inflammatory bowel disease macrophages. Clin Exp Immunol 2004; 136: 157–167.
Cao WJ, Li MH, Li JX, Xu X, Ren SX, Rajbanshi B, Xu JF . High expression of cathepsin E is associated with the severity of airflow limitation in patients with COPD. COPD 2016; 13: 160–166.
Rao NV, Rao GV, Hoidal JR . Human dipeptidyl-peptidase I. Gene characterization, localization, and expression. J Biol Chem 1997; 272: 10260–10265.
Nakajima R, Ishida M, Kamiya CA, Yoshimatsu J, Suzuki M, Hirota A, Ikeda T, Harigaya T . Elevated vasoinhibin derived from prolactin and cathepsin D activities in sera of patients with preeclampsia. Hypertens Res 2015; 38: 899–901.
Herías V, Biessen EA, Beckers C, Delsing D, Liao M, Daemen MJ, Pham CC, Heeneman S . Leukocyte cathepsin C deficiency attenuates atherosclerotic lesion progression by selective tuning of innate and adaptive immune responses. Arterioscler Thromb Vasc Biol 2015; 35: 79–86.
Pagano MB, Bartoli MA, Ennis TL, Mao D, Simmons PM, Thompson RW, Pham CT . Critical role of dipeptidyl peptidase I in neutrophil recruitment during the development of experimental abdominal aortic aneurysms. Proc Natl Acad Sci USA 2007; 104: 2855–2860.
Wolinsky H, Capron L, Goldfischer S, Capron F, Coltoff-Schiller B, Kasak LE . Hydrolase activities in the rat aorta. II. Effects of hypertension alone and in combination with diabetes mellitus. Circ Res 1978; 42: 831–839.
Acknowledgements
This study was supported by a grant from the National Institute of Health NICHD R21HD076289 to YW.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
About this article

Cite this article
Gu, Y., Lewis, D., Alexander, J. et al. Upregulation of cathepsin C expression contributes to endothelial chymase activation in preeclampsia. Hypertens Res 40, 976–981 (2017). https://doi.org/10.1038/hr.2017.73
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/hr.2017.73
Keywords
This article is cited by
-
Expression and significance of cathepsin C and cathepsin D during pregnancy and Preeclampsia
Reproductive Biology and Endocrinology (2023)
-
Histone deacetylase inhibition disturbs the balance between ACE and chymase expression in endothelial cells: a potential mechanism of chymase activation in preeclampsia
Hypertension Research (2019)
