
Abstract
Angiotensin II (AngII) is a crucial hormone that affects vasoconstriction and exerts hypertrophic effects on vascular smooth muscle cells. Here, we showed that phosphatidylinositol 3-kinase-dependent calcium mobilization plays pivotal roles in AngII-induced vascular constriction. Stimulation of rat aortic vascular smooth muscle cell (RASMC)-embedded collagen gel with AngII rapidly induced contraction. AngII-induced collagen gel contraction was blocked by pretreatment with a phosphatidylinositol 3-kinase (PI3K) inhibitor (LY294002) whereas ERK inhibitor (PD98059) was not effective. AngII-induced collagen gel contraction was significantly blocked by extracellular calcium depletion by EGTA or by nifedipine which is an L-type calcium channel blocker. In addition, AngII-induced calcium mobilization was also blocked by nifedipine and EGTA, whereas intracellular calcium store-depletion by thapsigargin was not effective. Finally, pretreatment of rat aortic ring with LY294002 and nifedipine significantly reduced AngII-induced constriction. Given these results, we suggest that PI3K-dependent activation of L-type calcium channels might be involved in AngII-induced vascular constriction.
Similar content being viewed by others
Introduction
The renin-angiotensin system (RAS) plays crucial roles in many physiological processes. Angiotensin II (AngII) is formed from a series of proteolytic cleavages of angiotensinogen to angiotensin I (AngI) by renin, and subsequent cleavage of AngI to AngII by angiotensin converting enzyme (ACE). Acute roles for AngII have been implicated in the regulation of plasma volume control, vasoconstriction, and blood pressure control, whereas chronic stimulation with AngII induces hyperplasia and hypertrophy of vascular smooth muscle cells (VSMCs) (Geisterfer et al., 1988).
Among its pleiotropic effects, vasoconstriction-induced blood pressure control is the major function of AngII. The receptors for AngII include angiotensin type 1 receptor (AT1R) and angiotensin type 2 receptor (AT2R), which belong to the seven trans-membrane superfamily that is coupled with G proteins. Occupation of AT1R by AngII results in intracellular calcium mobilization, which binds to calmodulin and activates myosin light chain kinase (MLCK). Activated MLCK eventually leads to the phosphorylation of the myosin light chain which causes it to interact with actin, causing contraction of VSMCs (Yan et al., 2003). MLCK is negatively regulated by myosin light chain phosphatase (MLCP), which is inactivated by Rho kinase. Activation of the Rho kinase signaling pathway leads to sustained contraction of VSMCs (Uehata et al., 1997). Therefore, intracellular calcium mobilization is a key mediator of VSMC contraction.
Binding of AngII to AT1R has been reported to activate several G proteins including Gαq/11, Gα12/13, and Gβγ complexes. Activation of G proteins leads to the activation of downstream signaling molecules such as PLC, PLD, and PLA2 (Ushio-Fukai et al., 1999a). Among the PLC isoforms, PLC-β isoform is coupled with G proteins and generates inositol tris-phosphate (IP3) from phosphatidylinositol 4,5-bisphosphate (PIP2) by hydrolytic cleavage (Rhee, 2001). IP3 evokes calcium mobilization by stimulating IP3 receptors on intracellular calcium storage sites such as the endoplasmic reticulum. Elevation of intracellular calcium concentrations eventually leads to VSMC contraction. In addition to the effect of AngII on calcium mobilization, AngII also causes membrane NAD(P)H oxidase to generate reactive oxygen species (ROS) such as superoxide and hydrogen peroxide (H2O2). For instance, AngII causes NAD(P)H oxidase subunits Nox1 and Nox4 to generate ROS (Lassegue et al., 2001), although the molecular mechanism is still not clear. AngII-dependent production of ROS affects the activation of many signaling molecules including p38 MPAK, Akt, and ERK1/2 (Ushio-Fukai et al., 1999b; Nishida et al., 2005). Although p38 MAPK and ERK1/2 are activated by AngII stimulation, these kinases are implicated in VSMC differentiation, proliferation and migration (Sugden and Clerk 1997; Taniyama et al., 2004).
AngII also cross-communicates with several tyrosine kinases, including receptor tyrosine kinases such as EGF-R, PDGF-R, insulin receptor (IR), and nonreceptor tyrosine kinases such as the c-Src family kinases, calcium-dependent proline-rich kinase 2 (Pyk2), focal adhesion kinase (FAK) and Janus kinase (JAK). The major function of AngII-dependent activation of nonreceptor tyrosine kinases seems to be the regulation of focal adhesion complex formation and actin bundling (Taniyama et al., 2003). AngII can trans-activate receptor tyrosine kinases independent of growth factors. For instance, stimulation of VSMC with AngII stimulates formation of Shc and Grb2 complex with PDGF-R (Linseman et al., 1995). Once activated, receptor tyrosine kinases evoke the phosphatidylinositol 3-kinase (PI3K)/3-phosphoinositide-dependent kinase-1 (PDK-1)/Akt cascade, which leads to growth, metabolism, survival, and remodeling of VSMCs.
The major physiological function of AngII is VSMC contraction and maintaining vascular tone through the regulation of calcium mobilization. Although PLC-β-dependent intracellular calcium mobilization during AngII stimulation is important for VSMC contractions, recent evidence also supports the idea that AngII increases myocardial activity through the augmentation of inward calcium currents through L-type calcium channels (Baker et al., 1992). Other evidence has supported the argument that Gβγ directly activates PI3Kγ isoform to generate PIP3 (Viard et al., 1999), which eventually stimulates L-type calcium channels in vascular myocytes from portal vein (Le Blanc et al., 2004). However, there is no direct evidence that PI3K-dependent calcium mobilization is important for vascular constriction upon AngII stimulation. Here we provide strong evidence that AngII-dependent rat aortic ring constriction is mediated by PI3K-mediated L-type calcium channel activation.
Results
AngII-induced contraction of isolated RASMCs
Previously, it was reported that AngII regulates calcium mobilization through PI3K-β-dependent L-type calcium channel activation in vascular myocytes from portal vein (Viard et al., 1999). To understand the role of PI3K-dependent L-type calcium channel modulation in VSMCs from thoracic aorta, we isolated rat aortic smooth muscle cells (RASMCs). As shown in Figure 1A, isolated RASMC expressed vascular smooth muscle cell marker proteins such as smooth muscle actin (SMA) and calponin. On the other hand, RASMCs did not express endothelial nitric oxide synthase (eNOS) which is an endothelial cell marker protein. A10 and human umbilical vein endothelial cell (HUVEC) extracts were used as positive controls of smooth muscle and endothelial cells, respectively. Isolated RASMCs rapidly contracted within 5 min after exposure to 1 µM of AngII (Figure 1B and 1C). However, AngII did not induce the contraction of A10 and HUVEC cells (data not shown).

AngII-induced RASMC contraction. (A) Cells from rat aortic smooth muscle were subjected to western blot analysis using the indicated antibodies. A10 and HUVEC cell lysates were included as markers for smooth muscle and endothelial cells, respectively. (B) Isolated RASMCs were embedded in collagen gel matrix as described in Methods. Gel contraction was initiated by addition of AngII (1 µM), and images were captured as described in Methods. Images are representative of two independent experiments each done in triplicate. (C) RASMC-embedded collagen gel was stimulated with either vehicle or AngII (1 µM), images were taken, and the surface area of the collagen gel was analyzed as described in Methods. Data are mean ± S.D. of two independent experiments (n = 3 for each experiment).
PI3K- and L-type calcium channel-dependent RASMC contraction
Since PI3K is involved in the AngII-induced calcium mobilization of smooth muscle cells from portal vein (Viard et al., 1999), we next examined the possible role of PI3K in VSMC contractions. As shown in Figure 2A, pretreatment with the PI3K inhibitor, LY294002 (10 µM), significantly reduced AngII-induced RASMC contractions. However, inhibition of the ERK1/2 signaling pathway by PD98059 (10 µM) slightly potentiated AngII-induced RASMC contractions. Inhibition of PLC by U73122 (10 µM) also blocked AngII-induced RASMC contraction (data not shown). Since calcium mobilization is important for smooth muscle cell contraction, we examined the effect of calcium channel blocker on AngII-induced RASMC contractions. As shown in Figure 2B, chelation of extracellular calcium by EGTA (5 mM) or blocking L-type calcium channels by nifedipine (20 µM) strongly inhibited AngII-induced RASMC contraction. However, depletion of intracellular calcium by thapsigargin (1 µM) did not affect AngII-induced RASMC contraction.

PI3K and L-type calcium channels are important for RASMC contraction in vitro. (A) RASMCs-embedded in collagen gel matrix were pretreated with vehicle, LY294002 (10 µM), or PD98059 (10 µM) for 20 min, and then stimulated with AngII (1 µM). Data are mean ± S.D. of two independent experiments (n = 3 for each experiment). (B) RASMCs-embedded in collagen gel matrix were pretreated with vehicle, EGTA (5 mM), thapsigargin (1 µM), or nifedipine (20 µM) for 20 min, and then stimulated with AngII (1 µM). Data are mean ± S.D. of two independent experiments (n = 3 for each experiment).
Inhibition of calcium mobilization by PI3K and L-type calcium channel inhibitors
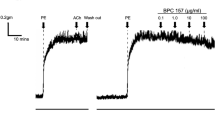
Since PI3K and L-type calcium channel inhibitors strongly blocked AngII-induced RASMC contraction, we examined the effect of these inhibitors on intracellular calcium mobilization. As shown in Figure 3A, pretreatment of RASMC with LY294002 completely blocked AngII-induced calcium mobilization, whereas pretreatment of PD98059 did not affect AngII-induced calcium mobilization. AngII-induced calcium mobilization was also completely blocked by extracellular calcium chelation (Figure 3B). Inhibition of L-type calcium channels by nifedipine significantly attenuated AngII-induced calcium mobilization.

AngII induces calcium mobilization through PI3K- in an L-type calcium channel-dependent manner. (A) RAMSCs were pretreated with vehicle, LY294002 (10 µM), or PD98059 (10 µM) for 20 min, followed by stimulation with AngII (1 µM). Images are representative of three independent experiments. The time of AngII stimulation is indicated by an arrow head. (B) RASMCs were pretreated with vehicle, thapsigargin (1 µM), EGTA (5 mM), or nifedipine (20 µM) for 20 min, followed by stimulation with AngII (1 µM). Images are representative of three independent experiments. The time of AngII stimulation is indicated by an arrow head.
PI3K- and L-type calcium channel-dependent constriction of rat aorta
Since calcium mobilization and contraction of RASMC were mediated by PI3K and L-type calcium channels, we extended the experiments to ex vivo conditions to clarify the effect of PI3K and L-type calcium channels on vascular constriction. As shown in Figure 4A, stimulation of thoracic aorta with AngII resulted in about 30-35% contraction of vessels. AngII-induced vascular contraction was significantly blocked by a PI3K inhibitor, LY294004, whereas inhibition of ERK1/2 signaling pathway did not affect AngII-induced vascular contractions. Depletion of extracellular calcium by EGTA strongly inhibited AngII-induced vascular contraction (Figure 4B). In addition, blocking of L-type calcium channels by nifedipine significantly inhibited AngII-induced vascular contractions. In contrast, depletion of intracellular calcium stores by thapsigargin did not affect AngII-induced vascular contraction.

PI3K and L-type calcium channels regulate AngII-induced contractions of thoracic aorta. (A) Thoracic aortic rings were isolated from rats and pretreated with vehicle alone, LY294002 (10 µM), or PD98059 (10 µM) for 20 min, and then stimulated with AngII (1 µM). Contraction is expressed as a percent of KCl (60 mM)-induced maximum tension. Data are mean ± S.D. of two independent experiments (n = 2 for each experiment). *Significantly different from vehicle-pretreated aortic rings (P < 0.05). **Values were not statistically different from vehicle-pretreated aortic rings (P > 0.05). (B) The AngII (1 µM)-induced tension of thoracic aortic rings was measured after pretreatment with vehicle, EGTA (5 mM), nifedipine (20 µM), or thapsigargin (1 µM) for 20 min.
Discussion
A plethora of evidence suggests that AngII plays key roles in the regulation of blood pressure. AngII acutely increases blood pressure by induction of blood vessel constriction. In fact, antagonists of AT1R effectively reduced blood pressure in hypertensive rats (Dendorfer et al., 2005). It has been reported that AngII activates many signaling pathways including the PLC, Ras/MEKK/MAPK, and p38 MAPK pathways (Smith et al., 1984; Meloche et al., 2000; Ishihata et al., 2002). In addition, AngII stimulates PI3K through the modulation of Gβγ (Viard et al., 1999). Likewise, pharmacological inhibition of PI3K resulted in the attenuation of RASMC contraction as well as in calcium mobilization (Figure 2-4). However, MAPK signaling pathway seems not to be involved in AngII-dependent physiological responses. Currently, the involvement of MAPK signaling pathways in VSMC contractions is controversial. Nevertheless, our results clearly showed that pharmacological inhibition of MAPK signaling pathway did not affect VSMC contraction both in vitro and ex vivo (Figure 2A and 4A). More importantly, inhibition of MAPK signaling pathway did not alter AngII-induced calcium mobilization (Figure 3A), which is a very important signaling response during AngII-induced vascular smooth muscle contraction. Therefore, the MAPK signaling pathway seems to be involved in prolonged responses such as proliferation and differentiation (Marrero et al., 1997; Yamada et al., 1999) rather than in acute responses.
AngII-induced calcium mobilization is a crucial event during VSMC contraction. Apparently, AngII stimulates PLC isoforms to generate IP3 through the activation of Gαq/11, G12/13, and Gβγ. Calcium is released from endoplasmic reticulum via IP3-operated calcium channels. Hence, PLC-mediated calcium mobilization has been implicated in the regulation of VSMC contraction. The possible involvement of PLC was suggested by experiments in which electroporation of anti-Gαq/11 antibody to VSMC suppressed AngII-induced IP3 production (Ushio-Fukai et al., 1998). However, there is no direct link between IP3-operated calcium mobilization and VSMC contraction. In contrast, our data showed that depletion of intracellular calcium stores by thapsigargine inhibited 50% of AngII-induced calcium mobilization (Figure 3B). This result indicates that AngII partially stimulates IP3-operated calcium mobilization. However, blocking IP3-operated calcium mobilization was not enough to block AngII-induced VSMC contractions both in vitro and ex vivo (Figure 2B and 4B). These results suggest that another mechanism of calcium mobilization is important for VSMC contractions. Currently, the involvement of PLC-mediated signaling pathways in VSMC contractions is still ambiguous. Studies using a genetically modified mouse model of PLC will provide detailed molecular mechanism of PLC-dependent VSMC contraction.
Although AngII-induced PLC activation and subsequent calcium mobilization have been implicated as major signaling pathways, it has been reported that AngII also induces the activation of growth factor receptor signaling pathways. For instance, stimulation of VSMC with AngII activates the tyrosine kinase signaling pathway including MAPK (Watts et al., 1998). However, pharmacological inhibition of MAPK does not affect VSMC contraction. Likewise, our results showed that pharmacological inhibition of the MAPK signaling pathway affected neither AngII-induced VSMC contraction nor calcium mobilization (Figure 2A, 3A and 4A). The involvement of other signaling pathways rather than G protein-coupled PLC activation has been implicated by recent studies. The atypical PI3K class is directly activated by the Gβγ subunits of G proteins (Viard et al., 1999). Consistent with this, our results showed that pharmacological inhibition of PI3K strongly attenuated AngII-induced calcium mobilization and VSMC contraction (Figure 3A and 4A). PI3K-dependent activation of calcium mobilization was further demonstrated by Le Blanc et al. (2004). They showed that PIP3, which is a product of PI3K, can directly activate L-type calcium channels. It is also supported by our results that depletion of extracellular calcium or blocking of L-type calcium channels significantly blocks AngII-induced calcium mobilization (Figure 3B). In addition, the link between L-type calcium channel opening and VSMC contraction was further confirmed by showing that inhibition of L-type calcium channels blunted AngII-induced VSMC contraction both in vitro and ex vivo (Figure 2B and 4B). Therefore, it is quite clear that direct activation of L-type calcium channels by PIP3 is important for the regulation of VSMC contraction. The downstream effector molecule of PIP3 seems not to be involved in AngII-dependent VSMC contractions since pharmacological inhibition of Akt/PKB by SH-5 (10 µM) did not affect AngII-induced VSMC contraction (data not shown).
Because inhibition of PLC activity by U73122 also suppressed AngII-induced VSMC contraction (data not shown) we cannot exclude possible involvement of PLC activation in AngII-dependent VSMC contraction. Consistent with this, depletion of intracellular calcium stores by thapsigargin resulted in a 50% reduction of AngII-induced calcium mobilization (Figure 3B), demonstrating PLC-mediated calcium mobilization in AngII-stimulated VSMCs. Nonetheless, pretreatment of thapsigargin barely affected AngII-induced VSMC contractions (Figure 4B). These results could be interpreted as follows. First, it is possible that depletion of intracellular calcium stores is not sufficient for the dropping effective concentration of calcium to evoke VSMC contraction. Second, in addition to calcium mobilization, another signaling pathway can be diverted from PLC, and lead the VSMC contraction. Consistent with this idea, it has been reported that PKC also regulates AngII-induced VSMC contractions (Yu et al., 2004). Therefore, multiple signaling pathways seem to be involved in the regulation of AngII-induced VSMC contractions.
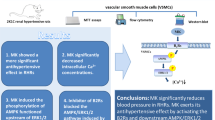
In summary, we have demonstrated that AngII-induced aortic ring constriction is mediated by PI3K-dependent regulation of L-type calcium channel activity. Our studies suggest that changes in intracellular calcium concentration by L-type calcium channels are important for arterial smooth muscle contraction, which is important for in vivo responses in AngII-dependent acute blood pressure regulation. The analysis of calcium modulation using knock-out animal models will shed more light on the AngII-mediated signaling pathway of VSMC contraction.
Methods
Reagents
All cell culture media and supplements were purchased from Cambrex Corp. Anti-SMA and anti-calponin antibodies were purchased from Sigma-Aldrich. Anti-β-actin antibody was obtained from MP Biomedicals. Anti-eNOS antibody was from BD Transduction Laboratories. Collagen gel was purchased from BD Bioscience and Fura-2-AM was from Invitrogen. LY294002 and PD98059 were obtained from Calbiochem. Nifedipine, thapsigargin, collagen, gelatin, and AngII were purchased from Sigma-Aldrich. All other high quality reagents were purchased from Sigma-Aldrich unless otherwise indicated.
Rat aortic smooth muscle cell (RASMC) preparation
Pathogen-free Sprague-Dawley rats (6 to 8 weeks old) were housed under diurnal lighting conditions and allowed unlimited access to feed and tap water. All animal experiments were performed under university animal welfare guidelines. Aortic smooth muscle cells were isolated from the thoracic aorta by a tissue explanting method as described previously (Hall et al., 1991). Briefly, sodium barbital anesthetized rats were perfused with PBS for 5 min. Thoracic aorta was aseptically isolated and surrounding fat and connective tissues were discarded. Vessels were longitudinally cut and the lumen side was scraped with a razor blade to remove the intima. Vessels were fragmented into 3-5 mm lengths and explanted lumen side down on collagen-coated culture dishes. Tissue fragments were maintained in DMEM/10% FBS with changing of the medium every two days. After seven days of explanting, tissue fragments were discarded and sprouted VSMCs were collected and used for experiments.
Collagen gel contraction assay
To measure RASMC contractility, RASMCs from confluent cultures were trypsinized, resuspended in serum free DMEM and diluted to 1 × 106 cells/ml. The cell suspension was mixed on ice with collagen gel solution (6 mg/ml of collagen type I in 2× PBS pH8.0) to give 5 × 105 cells/ml and 3 mg/ml of collagen gel solution. One hundred µl of RASMC-collagen gel solution was added per well to 24-well plates. Where indicated, various inhibitors were added. The plate was incubated at 37℃ to allow for gel polymerization. After 1 h, the gels were floated with serum-free DMEM and AngII was added to initiate contraction while images were captured using a digital charge-coupled device camera. Collagen gel contraction was measured as a decrease in gel area using Scion Image software (compliments of Scion Corporation, Frederick, MD, http://www.scioncorp.com). Relative gel area was obtained by dividing the area at each time point by the initial area of the gel.
Measurement of cytosolic calcium concentration
Intracellular calcium concentration was measured using fura-2/AM, a calcium-sensitive fluorescent dye, as described previously (Grynkiewicz et al., 1985). Briefly, a total of 2 × 106 RASMCs were incubated with 3 µM fura-2/AM at 37℃ in fresh serum-free DMEM medium with stirring for 50 min. A total of 2 × 105 cells were aliquoted for each assay into Locke's solution (154 mM NaCl, 5.6 mM KCl, 1.2 mM MgCl2, 5 mM HEPES, pH 7.3, 10 mM glucose, 2.2 mM CaCl2, and 0.2 mM EGTA). Fluorescence was measured at emission wavelength of 500 nm using an excitation wavelength of 340 nm and 380 nm.
Measurement of aortic ring tension
The thoracic aorta was rapidly removed and immersed in ice-cold modified Krebs' solution (118 mM of NaCl, 4.7 mM of KCl, 1.2 mM of MgSO4·7H2O, 1.2 mM of KH2PO4, 25 mM of NaHCO3, 2.5 mM of CaCl2·2H2O, 5.6 mM of glucose, pH 7.4). The aorta was dissected to remove fat and connective tissue and sliced into rings (2-3 mm in length). The aortic rings were suspended by two L-shaped stainless-steel wires inserted into the lumen followed by immersion in 10 ml organ chambers filled with Krebs' solution. The basal tension of the aortic ring was maintained at 2 g and aerated continuously with 95% O2/5% CO2. Changes in isometric tension were recorded using a force-displacement transducer (Grass FT 0.3, Quincy, MA) connected to a Power Lab system 400 (ML 118, PowerLab, AD Instruments, Medford, MA) and saved in a computer. The aortic rings were allowed to equilibrate for 90 min while changing the chamber solution every 15 min. After resting tension of each vascular specimen had stabilized, maximum tension of each aortic ring was measured after addition of 60 mM KCl. Constricted aortic rings were relaxed by washing with Krebs' solution for 2 h. Various inhibitors were added 20 min prior to AngII stimulation. AngII-induced contraction was expressed as percent of 60 mM KCl-induced contraction.
Western blot analysis
VSMCs were lysed in lysis buffer containing 20 mM Tris-HCl, pH 7.3, 1 mM EGTA/EDTA, 1% Triton X-100, 1 mM Na3VO4, 10 µg/ml leupeptin and 10 µg/ml aprotinin. After centrifugation at 12,000 rpm for 5 min, 30 µg/ml of total protein was loaded onto a 10% polyacrylamide gel and, after electrophoresis, transferred to nitrocellulose membranes. Membranes were incubated with the indicated primary antibodies and IR Dye-conjugated secondary antibody. Protein bands were visualized by an infrared image analyzer (Li-COR Bioscience).
Statistical analysis
Results are expressed as the mean ± S.D. of two independent experiments (n = 3 for each experiment). When two groups were compared, an unpaired Student's t-test was used to assess differences. P-values less than 0.05 were considered significant.
Abbreviations
- Akt/PKB:
-
Akt/protein kinase B
- AngII:
-
angiotensin II
- PI3K:
-
phosphatidylinositol 3-kinase
- PIP3:
-
phosphatidylinositol 3,4,5-trisphosphate
- RASMC:
-
rat aortic smooth muscle cell
- VSMC:
-
vascular smooth muscle cell
References
Baker KM, Booz GW, Dostal DE . Cardiac actions of angiotensin II: Role of an intracardiac renin-angiotensin system . Ann Rev Physiol 1992 ; 54 : 227 - 241
Dendorfer A, Dominiak P, Tempel K, Raasch W . Peripheral sympatholytic actions of four AT1 antagonists: are they relevant for long-term antihypertensive efficacy ? J Hypert 2005 ; 23 : 1861 - 1867
Geisterfer AA, Peach MJ, Owens GK . Angiotensin II induces hypertrophy, not hyperplasia, of cultured rat aortic smooth muscle cells . Circ Res 1988 ; 62 : 749 - 756
Grynkiewicz G, Poenie M, Tsien RY . A new generation of Ca2+ indicators with greatly improved fluorescence properties . J Biol Chem 1985 ; 260 : 3440 - 3450
Hall KL, Harding JW, Hosick HL . Isolation and characterization of clonal vascular smooth muscle cell lines from spontaneously hypertensive and normotensive rat aortas . In Vitro Cell Dev Biol 1991 ; 27A : 791 - 798
Ishihata A, Tasaki K, Katano Y . Involvement of p44/42 mitogen-activated protein kinases in regulating angiotensin II- and endothelin-1-induced contraction of rat thoracic aorta . Eur J Pharm 2002 ; 445 : 247 - 256
Lassegue B, Sorescu D, Szocs K, Yin Q, Akers M, Zhang Y, Grant SL, Lambeth JD, Griendling KK . Novel gp91(phox) homologues in vascular smooth muscle cells : nox1 mediates angiotensin II-induced superoxide formation and redox-sensitive signaling pathways . Circ Res 2001 ; 88 : 888 - 894
Le Blanc C, Mironneau C, Barbot C, Henaff M, Bondeva T, Wetzker R, Macrez N . Regulation of vascular L-type Ca2+ channels by phosphatidylinositol 3,4,5-trisphosphate . Circ Res 2004 ; 95 : 300 - 307
Linseman DA, Benjamin CW, Jones DA . Convergence of angiotensin II and platelet-derived growth factor receptor signaling cascades in vascular smooth muscle cells . J Biol Chem 1995 ; 270 : 12563 - 12568
Marrero MB, Schieffer B, Li B, Sun J, Harp JB, Ling BN . Role of Janus kinase/signal transducer and activator of transcription and mitogen-activated protein kinase cascades in angiotensin II- and platelet-derived growth factor-induced vascular smooth muscle cell proliferation . J Biol Chem 1997 ; 272 : 24684 - 24690
Meloche S, Landry J, Huot J, Houle F, Marceau F, Giasson E . p38 MAP kinase pathway regulates angiotensin II-induced contraction of rat vascular smooth muscle . Am J Physiol 2000 ; 279 : H741 - H751
Nishida M, Tanabe S, Maruyama Y, Mangmool S, Urayama K, Nagamatsu Y, Takagahara S, Turner JH, Kozasa T, Kobayashi H, Sato Y, Kawanishi T, Inoue R, Nagao T, Kurose H . Gα12/13- and reactive oxygen species-dependent activation of c-Jun NH2-terminal kinase and p38 mitogen-activated protein kinase by angiotensin receptor stimulation in rat neonatal cardiomyocytes . J Biol Chem 2005 ; 280 : 18434 - 18441
Rhee SG . Regulation of phosphoinositide-specific phospholipase C . Ann Rev Biochem 2001 ; 70 : 281 - 312
Smith JB, Smith L, Brown ER, Barnes D, Sabir MA, Davis JS, Farese RV . Angiotensin II rapidly increases phosphatidate-phosphoinositide synthesis and phosphoinositide hydrolysis and mobilizes intracellular calcium in cultured arterial muscle cells . Proc Natl Acad Sci USA 1984 ; 81 : 7812 - 7816
Sugden PH, Clerk A . Regulation of the ERK subgroup of MAP kinase cascades through G protein-coupled receptors . Cell Signal 1997 ; 9 : 337 - 351
Taniyama Y, Weber DS, Rocic P, Hilenski L, Akers ML, Park J, Hemmings BA, Alexander RW, Griendling KK . Pyk2- and Src-dependent tyrosine phosphorylation of PDK1 regulates focal adhesions . Mol Cell Biol 2003 ; 23 : 8019 - 8029
Taniyama Y, Ushio-Fukai M, Hitomi H, Rocic P, Kingsley MJ, Pfahnl C, Weber DS, Alexander RW, Griendling KK . Role of p38 MAPK and MAPKAPK-2 in angiotensin II-induced Akt activation in vascular smooth muscle cells . Am J Physiol Cell Physiol 2004 ; 287 : C494 - C499
Uehata M, Ishizaki T, Satoh H, Ono T, Kawahara T, Morishita T, Tamakawa H, Yamagami K, Inui J, Maekawa M, Narumiya S . Calcium sensitization of smooth muscle mediated by a Rho-associated protein kinase in hypertension . Nature 1997 ; 389 : 990 - 994
Ushio-Fukai M, Griendling KK, Akers M, Lyons PR, Alexander RW . Temporal dispersion of activation of phospholipase C-β1 and -γ isoforms by angiotensin II in vascular smooth muscle cells. Role of αq/11, α12, and βγ G protein subunits . J Biol Chem 1998 ; 273 : 19772 - 19777
Ushio-Fukai M, Alexander RW, Akers M, Lyons PR, Lassegue B, Griendling KK . Angiotensin II receptor coupling to phospholipase D is mediated by the betagamma subunits of heterotrimeric G proteins in vascular smooth muscle cells . Mol Pharm 1999a ; 55 : 142 - 149
Ushio-Fukai M, Alexander RW, Akers M, Yin Q, Fujio Y, Walsh K, Griendling KK . Reactive oxygen species mediate the activation of Akt/protein kinase B by angiotensin II in vascular smooth muscle cells . J Biol Chem 1999b ; 274 : 22699 - 22704
Viard P, Exner T, Maier U, Mironneau J, Nurnberg B, Macrez N . Gβγ dimers stimulate vascular L-type Ca2+ channels via phosphoinositide 3-kinase . FASEB J 1999 ; 13 : 685 - 694
Watts SW, Florian JA, Monroe KM . Dissociation of angiotensin II-stimulated activation of mitogen-activated protein kinase kinase from vascular contraction . J Pharm Exp Therap 1998 ; 286 : 1431 - 1438
Yamada H, Akishita M, Ito M, Tamura K, Daviet L, Lehtonen JY, Dzau VJ, Horiuchi M . AT2 receptor and vascular smooth muscle cell differentiation in vascular development . Hypert 1999 ; 33 : 1414 - 1419
Yan C, Kim D, Aizawa T, Berk BC . Functional interplay between angiotensin II and nitric oxide: cyclic GMP as a key mediator . Arterioscle Throm Vasc Biol 2003 ; 23 : 26 - 36
Yu J, Tokinaga Y, Ogawa K, Iwahashi S, Hatano Y . Sevoflurane inhibits angiotensin II-induced, protein kinase C-mediated but not Ca2+-elicited contraction of rat aortic smooth muscle . Anesthesiology 2004 ; 100 : 879 - 884
Acknowledgements
This work was supported for two years by Pusan National Research Grant to S.S.B.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Do, K., Kim, M., Kim, J. et al. Angiotensin II-induced aortic ring constriction is mediated by phosphatidylinositol 3-kinase/L-type calcium channel signaling pathway. Exp Mol Med 41, 569–576 (2009). https://doi.org/10.3858/emm.2009.41.8.062
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3858/emm.2009.41.8.062
Keywords
This article is cited by
-
New insights on mode of action of vasorelaxant activity of simvastatin
Inflammopharmacology (2023)
-
Increased AT1 receptor expression mediates vasoconstriction leading to hypertension in Snx1−/− mice
Hypertension Research (2021)
-
DL0805-2, a novel indazole derivative, relaxes angiotensin II-induced contractions of rat aortic rings by inhibiting Rho kinase and calcium fluxes
Acta Pharmacologica Sinica (2016)
-
The prolactin family hormones regulate vascular tone through NO and prostacyclin production in isolated rat aortic rings
Acta Pharmacologica Sinica (2015)
-
Guidelines for the Isolation and Characterization of Murine Vascular Smooth Muscle Cells. A Report from the International Society of Cardiovascular Translational Research
Journal of Cardiovascular Translational Research (2015)
