
Abstract
Many individuals with Parkinson’s disease (PD) develop cognitive deficits, and a phenotypic and molecular overlap between neurodegenerative diseases exists. We investigated the contribution of rare variants in seven genes of known relevance to dementias (β-amyloid precursor protein (APP), PSEN1/2, MAPT (microtubule-associated protein tau), fused in sarcoma (FUS), granulin (GRN) and TAR DNA-binding protein 43 (TDP-43)) to PD and PD plus dementia (PD+D) in a discovery sample of 376 individuals with PD and followed by the genotyping of 25 out of the 27 identified variants with a minor allele frequency <5% in 975 individuals with PD, 93 cases with Lewy body disease on neuropathological examination, 613 individuals with Alzheimer’s disease (AD), 182 cases with frontotemporal dementia and 1014 general population controls. Variants identified in APP were functionally followed up by Aβ mass spectrometry in transiently transfected HEK293 cells. PD+D cases harbored more rare variants across all the seven genes than PD individuals without dementia, and rare variants in APP were more common in PD cases overall than in either the AD cases or controls. When additional controls from publically available databases were added, one rare variant in APP (c.1795G>A(p.(E599K))) was significantly associated with the PD phenotype but was not found in either the PD cases or controls of an independent replication sample. One of the identified rare variants (c.2125G>A (p.(G709S))) shifted the Aβ spectrum from Aβ40 to Aβ39 and Aβ37. Although the precise mechanism remains to be elucidated, our data suggest a possible role for APP in modifying the PD phenotype as well as a general contribution of genetic factors to the development of dementia in individuals with PD.
Similar content being viewed by others

Introduction
Linkage analyses as well as genome-wide association and exome sequencing studies have uncovered at least 20 genes associated with idiopathic Parkinson’s disease (PD). Still, to date, the identified genes only explain a small portion of the genetic burden in PD. It is likely that genetic factors involved in bringing about a PD phenotype comprise both genetic variants of strong effect, which alone are causative, as well as variants of weaker effect, which contribute to disease risk or phenotypic modification.
A significant overlap between different neurodegenerative diseases has been described on the neuropathologic, the genetic and the phenotypic level.1, 2, 3, 4 Neuropathologically, the overlap is exemplified by the coexistence of hallmark features of both Alzheimer’s disease (AD) and PD in individuals with Lewy body disease.1 On the genetic level, common genetic variants in microtubule-associated protein tau (MAPT) represent risk factors for PD3, 4 whereas, at the same time, rare variants of strong effect in MAPT have long been recognized as a cause of frontotemporal dementia (FTD).2 Phenotypically, it is known that at least 30% of individuals with PD develop dementia5, 6 and that age has been described as a major predisposing factor for the development of cognitive impairment.7 Accordingly, we sought to assess the contribution of genetic factors known to be involved in dementias such as AD8, 9, 10, 11 or FTD2, 12, 13, 14 to the PD phenotype.
Methods
Standard protocol approvals, registrations and patient consents
Ethics review board approval was obtained at all participating institutions, with the primary review board located at the Technische Universität München, Munich, Germany. All the participants provided written informed consent for participation in the study.
Participants, variant screening and genotyping
We used Idaho LightScanner (BioFire Defense, Salt Lake City, UT, USA) melting curve analysis to screen the coding regions and exon–intron boundaries of β-amyloid precursor protein (APP), presenilin 1 and 2 (PSEN1 and PSEN2), tau (MAPT), TAR DNA-binding protein 43 (TDP-43), granulin (GRN) and fused in sarcoma (FUS) in 376 individuals with PD (188 with PD without dementia, 188 with PD plus dementia as diagnosed according to the guidelines set forth by the task force of the Movement Disorder Society15) and 376 KORA-AGE controls (APP and MAPT only; Supplementary Figure 1). In the case of altered melting patterns suggestive of variants, Sanger sequencing ensued.
Variants identified during the screening phase were genotyped in 975 PD cases, 93 independent neuropathologically confirmed cases of Lewy body disease, 613 AD, 182 FTD cases and 1014 controls using Sequenom MALDI-TOF mass spectrometry. For technical reasons, MAPT c.1637G>A (p.(R546H)) and PSEN2 c.211C>T (p.(R71W)) were not included. Two 3 base pair (bp) deletions in APP were assessed by fragment analysis as described previously.16 One variant (APP c.1795G>A (p.(E599K))) that showed significant association in the first sample was also assessed in a second independent sample of 715 PD cases and 948 healthy controls from Spain. Significance was judged using the χ2-test. For the genotyping experiments, P-values were corrected using the Bonferroni method. P-values given for burden tests represent nominal P-values. For a detailed description, see Supplementary Figure 1.
The following transcripts and genomic sequences were used in primer design and variant annotation: APP—NM_000484.3, NG_007376.1; PSEN1—NM_000021.3, NG_007386.2; PSEN2—NM_000447.2, NG_007381.1; FUS—NM_004960.3, NG_012889.2; GRN—NM_002087.2, NG_007886.1; MAPT—NM_001123066.3, NG_007398.1; TDP-43—NM_007375.3, NG_008734.1. Primer sequences are available upon request.
Immunohistochemistry
Cortical and midbrain sections of the individual harboring the APP c.1795G>A (p.(E599K)) variant were stained for Aβ and alpha-synuclein. Staining procedure and antibodies can be found in the supplement.
Cloning, transfections and analysis of Aß-spectrum
cDNA of the pCDNA3.1+APP695sw vector containing all identified APP variants were transiently transfected into HEK293 cells and Aβ was analyzed by mass spectrometry as depicted in the supplement in the culture medium.
Results
Variant screening of ‘dementia genes’ in individuals with PD
Within the coding regions and exon–intron boundaries (±10 bp) of APP, PSEN1, PSEN2, MAPT, FUS, TDP-43 and GRN, we identified a total of 27 rare variants with minor allele frequency (MAF) <5% in 376 individuals with PD (n=188; 70.4±11.73 years, 28.4% female) or PD+D (n=188; 72.0±6.1 years, 33.0% female). Interestingly, more individuals with PD+D (10.11%) than solely PD (4.26%) harbored a rare variant with MAF <5% in any of the seven ‘dementia genes’ (19 PD+D individuals with a variant vs 8 PD individuals with a variant; P=0.0027, χ2-test). Four individuals harbored the GRN c.1297C>T (p.(R433W)) (rs63750412) variant and one GRN c.103G>A (p.(G35R)). One novel variant in PSEN1 (c.442A>G (p.(I148W))) within two amino acids of variants known to affect the function as well as three previously reported variants in PSEN2 (c.185G>A (p.(R62H)) (rs58973334), c.211C>T (p.(R71W)) (rs140501902), c.389C>T (p.(S130L)) (rs63750197)) were found. No variants were identified in either TDP-43 or FUS. Nine were also found by the NHLBI-GO exome sequencing project.17 (Table 1) For a detailed discussion of the phenotype of variant carriers, please refer to the supplement. (Supplementary Table 1)
For APP and MAPT, the screening was performed in the above 376 PD cases and 376 KORA-AGE controls. In APP, 11 rare variants with MAF <5% (seven missense, two 3-bp deletions, two nearsplice variants) were seen. In total, 10 cases but only 4 general population controls carried a rare APP variant. None of these variants have previously been reported in individuals with a neurodegenerative condition. In MAPT, we identified a total of 10 rare variants (9 missense, 1 stop). Overall, seven cases and five controls harbored a rare MAPT variant. (Table 1,Supplementary Table 1). Analysis by common prediction algorithms yielded contradicting results for most variants (Table 1), thus warranting additional frequency assessment and functional study.
Frequency assessment in individuals with PD, AD and FTD
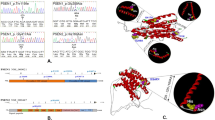
Frequency assessment for 25 of the 27 variants identified in the screening phase was carried out in a sample consisting of 975 PD patients (including the 376 used above), 613 AD patients, 182 FTD patients, 93 neuropathologically confirmed cases of Lewy body disease and 1014 controls (also including the 376 used above). 68.0% of the variants were very rare with MAF <0.1% in the control sample. When compared with controls, the APP c.1795G>A (p.(E599K)) variant was significantly more frequent in the PD phenotype than in controls (P=0.009, χ2-test; Supplementary Table 2) prior to correction for multiple testing. When publically available data from the NHLBI-ESP exomes17 (APP c.1795G>A (p.(E599K)) MAF=0.15% in KORA and 0.11% in NHLBI-ESP exomes vs 0.66% in PD cases) were added to the controls, the finding remained significant even after Bonferroni correction for multiple testing (14 out of 1068 cases vs 12 out of 5310 controls; Pnominal=3.8 × 10−7, Pcorrected=9.5 × 10−6, χ2-test). Exclusion of the 376 PD cases and 376 controls used in the discovery phase of the study did not alter this finding (11 out of 692 cases vs 11 out of 4934 controls; Pnominal=4.0 × 10−7, Pcorrected=1.0 × 10−5, χ2-test). However, when trying to replicate this finding in a Spanish PD case/control sample, we did not find any APP c.1795G>A (p.(E599K)) carriers in either cases or controls, possibly suggesting a population-specific effect of APP c.1795G>A (p.(E599K)) in Central Europeans. APP c.1795G>A (p.(E599K)) was the only variant identified in the 93 Lewy body disease cases. Neuropathologically, this case was indistinguishable from other LBD cases and showed no obvious special pathology. Clinically, this individual had suffered from classical, levodopa-responsive PD with an age of onset at 59 years. Her mother had also had PD. Histology revealed both Lewy bodies in the substantia nigra (SN) and some amyloid plaques in the frontal and parietal cortex and the hippocampus, in line with a diagnosis of idiopathic PD (Figure 1).

Location of rare variants in APP, GRN, MAPT, PSEN1, PSEN2 and histological features of an individual harboring the c.1795G>A (p.(E599K)) variant of APP. (a) Variants with MAF <5% found in PD cases are depicted above the schematic illustration of each gene, those found in controls – if the gene was analyzed in controls – below the gene. If variants were present more than once in the discovery sample, the number of occurrences is given in parentheses. Domain annotations were taken from Uniprot (accessed 12 December, 2012). HeP, heparin; AA, amino acids. (b) Depiction of a classical nigral Lewy body (left, antibody: anti-alpha-synuclein KM51, 1:1000, Novocastra/NCL-ASYN, counter stain: hematoxylin–eosine) and cortical Aβ plaques (right, antibody: 4G8, 1:2000, Signet) found in an individual with classical idiopathic PD and the APP c.1795G>A (p.(E599K)) variant. The neuropathology was in line with a cases of Lewy body disease (Braak stage 6) with additional Alzheimer-associated alterations (Braak and Braak Stage II), cerebral amyloid angiopathy (Thal stage 1) and beginning argyrophillic grain disease.
Burden tests analyzing the load of rare variants were performed for both APP and MAPT. This revealed an excess of rare variants with MAF <5% in APP in PD (27 individuals with a variant out of 975) when compared with either controls alone (13 out of 1014, P=0.018, χ2-test), AD cases (4 out of 613, P=0.002, χ2-test) or the combined sample of controls, AD and FTD cases (P=2.22 × 10−4, χ2-test). This excess of variants with MAF <5% in APP in PD (17 individuals with a variant out of 599) was also seen after exclusion of the ‘discovery’ samples when compared with the joined sample of controls, AD and FTD cases (14 out of 1433, Pcorrected=0.014, χ2-test) and to AD cases alone (4 out of 613, Pcorrected=0.014, χ2-test). When compared with the controls only (9 out of 638), variants were nearly twice as frequent (MAFPD=1.41% vs MAFKORA=0.71%) but this result fell short of statistical significance (Pcorrected=0.24, χ2-test). The frequency of rare variants in MAPT was similar in all the groups and remained unchanged after the omission of the initial 376 PD cases and 376 controls.
Impact of rare variants in APP on Aβ processing
Aβ spectral analysis was performed to further evaluate a potential functional effect of the identified coding variants in APP. In all but one, the Aβ spectrum reflected the wild-type situation. However, APP c.2125G>A (p.(G709S)), located within the Aβ domain, shifted the spectrum from Aβ40 as the main species to Aβ39 and – to a lesser extent – Aβ37 (Figure 2, Supplementary Figure 2).

Functional effect of rare APP variants. When overexpressed in HEK293 cells, the APP c.2125 (p.(G709S)) variant shifts the Aβ spectrum from Aβ40 to Aβ39 and Aβ37, whereas none of the other rare APP variants examined showed alterations in the Aβ spectrum.
Discussion
Screening of seven genes known to be strong genetic factors in AD or FTD in a sample comprising both individuals with PD and PD+D revealed a number of rare variants not previously described. Interestingly, identified variants in APP were more common in PD with and without dementia than in either controls or AD. Next, to a mere chance occurrence, there are several possible explanations for this finding. For one, rare variants in known dementia genes could represent phenotype modifiers in PD. This is supported by the fact that in the screening sample, rare variants were more frequent in the PD+D group than in the PD group when all seven genes were analyzed together. Also, the ‘dementia gene’ variants could contribute to the overall ‘neurodegenerative burden’, which reflects an increased susceptibility for neurodegenerative conditions in general. In this scenario, an excess of genetic alterations in a specific pathway plus additional non-genetic factors could then tip the balance toward one neurodegenerative phenotype or the other or create phenotypes in which features of multiple neurodegenerative diseases and symptoms coexist. Alternatively, this could also mean that the phenotypic spectrum of AD or FTD is broader than previously recognized and could include PD-like aspects.
The boldest proposal would be that rare variants of strong effect in APP or MAPT alone could cause PD. Mapt−/− mice have recently been shown to develop not only memory deficits but also PD-specific features such as a loss of neurons in the SN and reduced locomotion.18 Common variants in MAPT are an established risk factor for PD3, 4 and the relevance of allelic series – that is, both common variants of weak effect and rare variants of strong effect in one gene – to PD has already be shown.4, 19 Yet, in our sample, rare variants in APP, not MAPT, were enriched in PD. However, since a physical interaction between MAPT and APP and a role of MAPT in trafficking APP to the cell membrane has been reported,18, 20 rare variants in APP could have a similar effect with regard to PD as MAPT variants.
One of the identified APP variants (c.2125G>A (p.(G709S))) shifts the Aβ proteome spectrum from Aβ40 to Aβ39 and Aβ37 indicating that it likely interferes with γ-secretase cleavage. This could possibly be due to an alteration in the site at which APP interacts with γ-secretases, a mechanism recently postulated for increased Aβ37 production in response to an artificial APP variant (c.2095A>G (p.(K699E))) 10 amino acids N-terminal of our variant.21 None of the other APP variants showed an altered Aβ spectrum. However, further studies are necessary to exclude that these variants could affect the structure and, accordingly, the aggregation potential of generated Aβ as has been demonstrated for some AD-linked variants (reviewed in Haass et al22).
Yet, from our data we cannot conclude that an Aβ-related function is truly relevant to a potential (modifying) role in PD. Next to the well-recognized role in amyloid production, recently several other functions have been identified.23, 24 APP has been described to serve as a neuronal ferroxidase, which oxidizes Fe2+ and loads Fe3+ on to the iron transport protein transferrin.24 Moreover, iron accumulates in mice lacking App.24 As iron accumulation in the SN is a known feature of PD,25 it would be imaginable that APP dysfunction could also predispose to increased iron accumulation in the SN. App−/− mice also show increased cerebral levels of dopamine and catecholamines owing to a lack of amine catabolism via the amine oxidase function of App.23 Increased APP expression due to APP variants potentially related to PD could lead to cerebral dopamine deficits and a PD phenotype. Accordingly, APP’s ferroxidase24 and amine oxidase23 activities could even more plausibly fit a potential role in PD pathogenesis or phenotype modification and should be explored further.
References
Deramecourt V, Bombois S, Maurage CA et al: Biochemical staging of synucleinopathy and amyloid deposition in dementia with Lewy bodies. J Neuropathol Exp Neurol 2006; 65: 278–288.
Hutton M, Lendon CL, Rizzu P et al: Association of missense and 5'-splice-site mutations in tau with the inherited dementia FTDP-17. Nature 1998; 393: 702–705.
Pastor P, Ezquerra M, Munoz E et al: Significant association between the tau gene A0/A0 genotype and Parkinson's disease. Ann Neurol 2000; 47: 242–245.
Simon-Sanchez J, Schulte C, Bras JM et al: Genome-wide association study reveals genetic risk underlying Parkinson's disease. Nat Genet 2009; 41: 1308–1312.
Aarsland D, Andersen K, Larsen JP, Lolk A, Kragh-Sorensen P : Prevalence and characteristics of dementia in Parkinson disease: an 8-year prospective study. Arch Neurol 2003; 60: 387–392.
Riedel O, Klotsche J, Spottke A et al: Cognitive impairment in 873 patients with idiopathic Parkinson's disease. Results from the German Study on Epidemiology of Parkinson's Disease with Dementia (GEPAD). J Neurol 2008; 255: 255–264.
Kempster PA, O'Sullivan SS, Holton JL, Revesz T, Lees AJ. : Relationships between age and late progression of Parkinson's disease: a clinico-pathological study. Brain 2010; 133: 1755–1762.
Goate A, Chartier-Harlin MC, Mullan M et al: Segregation of a missense mutation in the amyloid precursor protein gene with familial Alzheimer's disease. Nature 1991; 349: 704–706.
Levy-Lahad E, Wasco W, Poorkaj P et al: Candidate gene for the chromosome 1 familial Alzheimer's disease locus. Science 1995; 269: 973–977.
Rogaev EI, Sherrington R, Rogaeva EA et al: Familial Alzheimer's disease in kindreds with missense mutations in a gene on chromosome 1 related to the Alzheimer's disease type 3 gene. Nature 1995; 376: 775–778.
Sherrington R, Rogaev EI, Liang Y et al: Cloning of a gene bearing missense mutations in early-onset familial Alzheimer's disease. Nature 1995; 375: 754–760.
Cruts M, Gijselinck I, van der Zee J et al: Null mutations in progranulin cause ubiquitin-positive frontotemporal dementia linked to chromosome 17q21. Nature 2006; 442: 920–924.
Neumann M, Rademakers R, Roeber S, Baker M, Kretzschmar HA, Mackenzie IR. : A new subtype of frontotemporal lobar degeneration with FUS pathology. Brain 2009; 132: 2922–2931.
Neumann M, Sampathu DM, Kwong LK et al: Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science 2006; 314: 130–133.
Emre M, Aarsland D, Brown R et al: Clinical diagnostic criteria for dementia associated with Parkinson's disease. Mov Disord 2007; 22: 1689–1707, quiz 1837.
Winkelmann J, Czamara D, Schormair B et al: Genome-wide association study identifies novel restless legs syndrome susceptibility loci on 2p14 and 16q12.1. PLoS Genet 2011; 7: e1002171.
(ESP) NGESP. Exome Variant Server. Available at http://evs.gs.washington.edu/EVS/ Accessed on December 1, 2012.
Lei P, Ayton S, Finkelstein DI et al: Tau deficiency induces parkinsonism with dementia by impairing APP-mediated iron export. Nat Med 2012; 18: 291–295.
Ross OA, Soto-Ortolaza AI, Heckman MG et al: Association of LRRK2 exonic variants with susceptibility to Parkinson's disease: a case-control study. Lancet Neurol 2011; 10: 898–908.
Guo JP, Arai T, Miklossy J, McGeer PL : Abeta and tau form soluble complexes that may promote self aggregation of both into the insoluble forms observed in Alzheimer's disease. Proc Natl Acad Sci USA 2006; 103:1953–1958.
Ousson S, Saric A, Baguet A et al: Substrate determinants in the C99 juxtamembrane domains differentially affect gamma-secretase cleavage specificity and modulator pharmacology. J Neurochem 2013; 125: 610–619.
Haass C, Kaether C, Thinakaran G, Sisodia S : Trafficking and proteolytic processing of APP. Cold Spring Harb Perspect Med 2012; 2: a006270.
Duce JA, Ayton S, Miller AA et al: Amine oxidase activity of beta-amyloid precursor protein modulates systemic and local catecholamine levels. Mol Psychiatry 2013; 18: 245–254.
Duce JA, Tsatsanis A, Cater MA et al: Iron-export ferroxidase activity of beta-amyloid precursor protein is inhibited by zinc in Alzheimer's disease. Cell 2010; 142: 857–867.
Dexter DT, Wells FR, Agid F et al: Increased nigral iron content in postmortem parkinsonian brain. Lancet 1987; 2: 1219–1220.
Acknowledgements
We are very grateful to Jelena Golic, Susanne Lindhof, Katja Junghans, Regina Feldmann and Sybille Frischholz at the Institute for Human Genetics at the Helmholtz Zentrum München for expert technical assistance.
All variants were submitted to dbSNP and received the NCBI numbers ss1399967350 to ss1399967376.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
This study was funded exclusively by in-house institutional funding from the Technische Universität München and the Helmholtz Zentrum München, Munich, Germany. There was no industry sponsorship. The KORA-Age project was funded by the German Ministry for Education and Research (BMBF) FKZ 01ET0713 and 01ET1003. BM has received grants from TEVA-Pharma, Desitin, Boehringer-Ingelheim and GE Healthcare and honoraria for consultancy from Bayer Schering Pharma AG and for presentations from GlaxoSmithKline and Orion Pharma as well as travel and meeting expenses from Boehringer-Ingelheim and Novartis. DH is currently employed by the Medical University of Vienna, Austria, and received research support through the National Institute of Neurological Disorders and Stroke Intramural Research Program and the Austrian Science Fund (FWF). He serves as member of the Medical Advisory Board of the International Essential Tremor Foundation and received honoraria and conference support from Ipsen and UCB. Walter Pirker received speaker honoraria from AOP Orphan Pharma, Medtronic Inc., Novartis, Boehringer-Ingelheim, Abbott Pharm and UCB as well as travel compensation from Ipsen Pharma, Boehringer-Ingelheim and Medtronic, Inc. BB and MJM receive research support from the Hungarian National Innovation Office (TÁMOP-4-2-1/B-03/1/KMR-2010-001). The functional analyses were supported by the European Research Council under the European Union's Seventh Framework Program (FP7/20072013)/ ERC Grant Agreement No. 321366-Amyloid (advanced grant to CH) and the general legacy of Mrs Ammer (to the Ludwig-Maximilians University/the chair of C.H.). PP is funded by a grant of the Spanish Ministry of Science and Innovation SAF-2010-22329-C02-01: 2011–2013. XE is funded by the Spanish Plan Nacional SAF-2008-00357 (NOVADIS); the Generalitat de Catalunya AGAUR 2009 SGR-1502; and the European Commission 7th Framework Program, Project No 261123 (GEUVADIS) and Project No 262055 (ESGI). JW serves on a scientific advisory board for UCB and has received speaker honoraria from UCB and Vifor Pharma. The remaining authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on European Journal of Human Genetics website
Supplementary information
Rights and permissions
About this article

Cite this article
Schulte, E., Fukumori, A., Mollenhauer, B. et al. Rare variants in β-Amyloid precursor protein (APP) and Parkinson’s disease. Eur J Hum Genet 23, 1328–1333 (2015). https://doi.org/10.1038/ejhg.2014.300
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ejhg.2014.300
This article is cited by
-
A thermodynamic investigation of amyloid precursor protein processing by human γ-secretase
Communications Biology (2022)
-
Genetic landscape of early-onset dementia in Hungary
Neurological Sciences (2022)
-
Reduced erythrocytic CHCHD2 mRNA is associated with brain pathology of Parkinson’s disease
Acta Neuropathologica Communications (2021)
-
Whole-exome sequencing of Finnish patients with vascular cognitive impairment
European Journal of Human Genetics (2021)
-
Singular cases of Alzheimer’s disease disclose new and old genetic “acquaintances”
Neurological Sciences (2021)
