
Abstract
The majority of genetic studies on Jewish populations have been focused on Ashkenazim, and genetic data from the Sephardic original source, the Iberian Peninsula, are particularly scarce. Regarding the mitochondrial genome, the available information is limited to a single Portuguese village, Belmonte, where just two different lineages (a single one corresponding to 93.3%) were found in 30 individuals. Aiming at disclosing the ancestral maternal background of the Portuguese Jewry, we enlarged the sampling to other crypto-Jewish descendants in the Bragança district (NE Portugal). Fifty-seven complete mtDNA genomes were newly sequenced and — in contrast with Belmonte — a high level of diversity was found, with five haplogroups (HV0b, N1, T2b11, T2e and U2e) being putatively identified as Sephardic founding lineages. Therefore — in sharp contrast with Belmonte — these communities have managed to escape the expected inbreeding effects caused by centuries of religious repression and have kept a significant proportion of the Sephardic founder gene pool. This deeper analysis of the surviving Sephardic maternal lineages allowed a much more comprehensive and detailed perspective on the origins and survival of the Sephardic genetic heritage. In line with previously published results on Sephardic paternal lineages, our findings also show a surprising resistance to the erosion of genetic diversity in the maternal lineages.
Similar content being viewed by others

Introduction
Sepharad, the traditional Jewish word for the Iberian Peninsula, became the name of the branch of the Jewish people who can trace their origin from there.1 Their settlement in Iberia certainly occurred a long time ago, and the oldest archeological evidence found so far has a chronology of 390 CE (http://www.uni-jena.de/en/News/PM120525_Schrifttafel.html). Thus, the Jewish presence in Portugal precedes the nation’s foundation in 1139 CE,2 and several documents for the period 1279–1325 testify the presence of Jewish communes in the Bragança district, NE Portugal.3 The degree of tolerance toward these communities was variable during the subsequent years and, in the sixteenth century, Iberian Jews were a demographically non-negligible minority with very heterogeneous social status. However, during that period (first in the United Kingdoms of Castile and Aragon under the rule of Catholic Kings and, shortly after, also in Portugal), Jews were forced into either conversion or expulsion. The Portuguese edict of expulsion was, however, far different from the one issued in Spain. As Jewish permanence in the country was intended, it was followed by several contradictory measures, as the forced baptism of 20 000 Jews from all over the country, who were prepared for exile and the forbiddance of inquiries on religion, during 20 years, for the New-Christians; hence, a consented crypto-Judaism (the secret adherence to Judaism while publicly professing another faith) was installed.2, 4, 5 Although a decree to end the distinction between Old and New-Christians was issued in 1507, the inquisition was established in 1536 and the effective abolishment of the distinction would only happen three centuries later with the Pombaline law.2
As the inquisition mainly targeted crypto-Judaism, it was especially rampant in the most remote areas of Portugal, near the Spanish border, such as Bragança and Belmonte. The persecutions reached Bragança region, Trás-os-Montes, in 1582–1583 causing numerous accusations and arrests from the very beginning.6, 7, 8 In the seventeenth and eighteenth centuries, the inquisitorial processes intensified and, as a result, there was a significant exodus to other countries, particularly of manufacturers and the merchant elite,9 many of them preserving connections with their original communities for a long period.7 The Jewish community of Bragança reappeared in the early twentieth century, gathering several families from the region, who had maintained their culture and religious secret practices for centuries. The estimated number of crypto-Jews at that time was around 700 to 800 people.7 Although the community was dissolved shortly after, a strong sense of belonging is still well alive today among the Jewish descendants.
Contemporary Jewish communities have been genetically analyzed both from population genetics and medical perspectives by means of uniparental and recombining markers,10, 11, 12, 13, 14, 15, 16, 17 and more recently also through genome-wide approaches.18, 19, 20, 21, 22, 23 However, only a few reports have been published on Sephardic and crypto-Jewish descendants.24, 25, 26, 27, 28, 29, 30
Recent analysis of paternal lineages in Iberia points to a high proportion of Jewish ancestry.10 Nonetheless, this work assumes an oversimplified parental population scenario and recognizes ‘alternative possible sources for lineages ascribed a Sephardic Jewish origin’.
Concerning Portugal, little information exists. For the Y chromosome, samples from Belmonte were included in the study from Adams et al10 but were pooled with non-Iberian Sephardic Jews; Nogueiro et al27 found an unexpected high haplotype diversity for an isolated, small-size population, scattered over the Bragança district (NE Portugal). Concerning mtDNA, previous studies identified some lineages with signatures of Sephardic origins found in Jewish communities from Portugal (Belmonte), Bulgaria, Italy, Turkey, Yugoslavia, Greece, Netherlands, Surinam and Spain12 and, more recently, South Texas, USA, and Mexico.31, 32 Nevertheless, data from the Iberian Peninsula, which constitutes the original geographic source of these populations, can be considered scarce: in Portugal, the information available until now was restricted to a single village, where just two different mtDNA lineages were found.12 Here, we are deepening preliminary results on control region (CR)33 to obtain a better picture of the Portuguese Jewish maternal lineages (Figure 1), in order to scrutinize whether the low diversity found in Belmonte is indeed a general hallmark of their mtDNA pool.

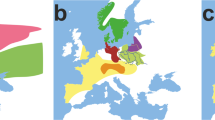
Geographic location of the sampling area in the Iberian Peninsula context. The zoomed area in the Iberian Peninsula represents the Portuguese district of Bragança and the Spanish province of Zamora (bold letter). Grey dots indicate the location of the sampling municipalities (uppercase) and villages (lowercase).
Materials and methods
Population sampling and DNA analyses
We sequenced complete mtDNA genomes from 57 unrelated self-designated Jews (and recognized as such by the host community) from the Bragança district, the same samples already characterized for Y chromosome27 and the mtDNA CR.33 Sampling criteria and collection method, as well as DNA extraction, are described in Nogueiro et al.27 The study was approved by the Ethics Committee of the University of Porto (N°02/CEUP/2012) and appropriate informed consent was required from all subjects. Full mtDNA sequences were obtained using the protocol described in Ramos et al.34, 35 Sequences were aligned against the revised Cambridge Reference Sequence GenBank accession number NC_012920.136 using Genious software version 5.5.8 (http://www.geneious.com), variants were annotated following the HGVS rules (http://www.hgvs.org/mutnomen/). Haplogroups were classified following the updated mtDNA phylogeny, PhyloTree, mtDNA tree Build 1637 (http://www.phylotree.org/) and assigned haplotypes were submitted to the EMPOP database38 (http://empop.org/). The accession number for the sequences reported in this paper is EMP00619.
Data analysis
Based on the mtDNA haplotypes found, standard and molecular diversity indices were estimated using ARLEQUIN software v3.5.1.3.39 For comparative purposes, original and publicly available data for mtDNA HVRI and HVRII (16 024–16 365 bp and 72–300 bp, respectively) were also compiled for 766 Jewish samples12 and 884 Western Iberian non-Jews.40, 41, 42
Differences in haplogroup composition, defined according to CR polymorphisms (16 024–16 400 bp for HVRI, and 073–340 bp for HVRII), among the analyzed Jews from Bragança and a large sample of the Portuguese host population,40 were assessed through Fisher’s exact tests implemented in IBM SPSS software.43
Median-joining networks44 (http://www.fluxus-engineering.com/sharenet.htm) of the (16 024–16 400 bp for HVRI, and 073–340 bp for HVRII) were constructed, for comparative purposes, using a compilation of original data on the Portuguese Jewish communities with publicly available data sets on the Portuguese host population,40 Mirandese41 and NW Spain.41, 42
Complementarily Median-joining networks using complete sequences were initially constructed and drawn, and then — for better visualization — redrawn for the defined putative Jewish founding lineages together with available complete sequences from public databases NCBI-GeneBank (http://www.ncbi.nlm.nih.gov/genbank/), empop.org (http://empop.org/), mtDNA community (http://www.mtdnacommunity.org/) and from Zhang et al45 work.
Results and Discussion
Haplotype diversity
Using complete mtDNA sequences, we were able to identify 45 haplotypes and 32 different haplogroups; detailed information is shown in Supplementary Table 1. We estimated global haplotype and nucleotide diversity to be just slightly lower than in the Portuguese host population, and much higher than in the Portuguese Jewish community from Belmonte (Table 1). Theta k values (θk) were also calculated for both the complete mtDNA as well as for the CR (Table 1). As expected, θk obtained for the CR is lower than when calculated for the complete mtDNA sequences, emphasizing the importance of studying complete mtDNA sequences in order to obtain more precise information on the maternal lineage diversity, as well as a more accurate estimate of the female effective population size.
Estimated θk value for the total sample is similar to the one usually found in Sephardic Jews from other countries, but exceptionally high if compared with Belmonte (Table 1), and also similar to the non-Jews from the neighboring region of Miranda.41 It is worth mentioning that the estimated number of putative female founders in each village of the Bragança district is higher than the current number of Jewish families. In fact, the samples included in the present work represent, if not the whole, the vast majority of the extant lineages, as increasing the sample size to reach predicted levels of saturation46, 47 would lead to the inclusion of closely related individuals.
The CR haplogroup frequencies of the Jews from Bragança together with the Portuguese host population and other geographical neighbors (Miranda41 and NW Spain41, 42) are presented in Supplementary Table 2. Differences in haplogroup composition were assessed through Fisher’s exact test among the Jews from Bragança and the Portuguese populations40 and Mairal et al.41 We focused on Jewish haplogroups that showed higher frequencies when compared with the Portuguese population, as they are likely to represent Sephardic origins (in accordance with Behar et al).12 Statistically significant differences were found for haplogroups HV0b (12.3% vs 0.0%; P=0.000), N1 (7.0% vs 1.3%; P=0.014), T2b11 (7.0% vs 0.0%; P=0.000), T2e (5.3% vs 0.7%; P=0.019) and U2e (7.0T vs 1.3%; P=0.014). These putative Jewish founding lineages account for 38.6% of the total sampled gene pool.
Sephardic lineages
The complex history of Jewish populations along the Mediterranean area is reflected on the haplogroup diversity observed in extant populations. An overview of their genetic composition is shown in Figure 2.

mtDNA haplogroup composition of the Sephardic Portuguese Jews (Bragança and Belmonte) and other Sephardic and non-Ashkenazi Jews. Haplogroups were defined for mtDNA HVRI and HVRII, 16 024–16 365 bp and 72–300 bp, respectively, according to Phylotree mtDNA tree Build 16 (other Sephardic and non-Ashkenazi Jews represent the same population as Table 1).
Until recently, haplogroup HV0, the ancestor of HV0b, was thought to have originated in Eastern Europe soon after the Last Glacial Maximum, having afterwards spread from there, following an east-west axis throughout Europe.48 However, the low gene and nucleotide diversities found in this region and in Northern Africa, compared with the ones found in North-Central Europe, seem to exclude Eastern Europe as a possible focus of expansion.49, 50
In the Iberian Peninsula, HV0 is a rare haplogroup, found at low frequencies in NW Spain, Zamora (4.7%),51 a Spanish province geographically close to Bragança. Because of a lack of resolution, it is not possible to differentiate haplogroup V inside HV0 in the Portuguese population.40 The same happens with the study of Mairal et al41 focused on a linguistically isolated population, Miranda do Douro, from the same geographical region (NE Portugal). Nevertheless, and considering the whole HV0 branch (haplogroup V included), we observe a frequency of 14% for this haplogroup, higher than the frequencies found in the Portuguese and in the Miranda populations (5.2% and 8.3%, respectively). Moreover, although HV0 has very low frequencies in eastern European Jewish Ashkenazim,52 we found a high prevalence of this lineage within the Jewish community of Bragança. Our results are in accordance with a previous report12 describing HV0 as a Jewish founding lineage in Portugal, as in Belmonte 93.3% of the analyzed mtDNA genomes could be traced back to a single female, carrying an mtDNA within haplogroup HV0b. Considering solely the CR, in a comparison between samples from Bragança Jews (present work), Portugal,40 Miranda41 and NW Spain,41, 42 our sample does not share any haplotypes with the Portuguese population (Supplementary Figure 1).
The most parsimonious tree of HV0b sub-clade, including all available mitochondrial complete sequences (Supplementary Figure S2), shows that Bragança HV0b lineage shares a common private variant m.8520A>G with Belmonte.12 Moreover, the Bragança samples cluster together, sharing a more recent variant not previously described m.10644G>A, which seems to have arisen locally. As no ethnic information is available for the remaining HV0b samples, and no introgression seems to have occurred with the Portuguese host-population, our results support the hypothesis that at least the HV0b-8520G haplotype is a Sephardic Jewish founding lineage.
The high frequency of haplogroup N1 found within the Jewish descendants of Bragança (7%) differentiates this community from the general Portuguese population, which displays an average frequency of just 0.8%. Haplogroup N1, along with macrohaplogroups M and R, is one of the founder lineages of the Eurasian settlement around 50–63 kya53 and comprises two major clades, N1a and N1b. N1a dates to approximately 20 kya and is a relatively rare haplogroup in Europe, reaching higher frequencies in Eastern Africa and in the Arabian Peninsula.54 Although this haplogroup was found in Neolithic and Mesolithic skeletal remains from Central and Northwest Europe,55, 56, 57 it was absent in previous studies on Iberian Neolithic populations, suggesting a rather uneven genetic and geographical spread of this lineage across Europe.58, 59 However, more recently, Gamba et al60 detected this lineage in Neolithic northeast Iberia, evidencing a possible scenario of several Neolithic colonization events along the Mediterranean, from the Near East. The samples from Bragança cluster into sub-haplogroups N1a and N1b and similarly to what was observed for HV0, NJ networks of N1b revealed no CR haplotype sharing with the general Portuguese population, except for four individuals from Miranda, an isolated population from the same geographical area as well as with two samples from Zamora41, 42 (Supplementary Figure 3).
Within N1b, the N1b2 haplogroup is considered as a founding lineage in Ashkenazi Jews with a ‘Hebrew/Levantine’ origin.61 However, an alternative European origin encompassing an assimilation of this lineage into the founding Ashkenazi population along the north Mediterranean coast has been recently proposed.52 Unexpectedly, we found no samples belonging to the N1b2 haplogroup among the Bragança Jews — all N1b samples fall inside the N1b1 sister-clade (N1b1a2 and N1b1a5) and the N1a1a1a2 sub-haplogroup. According to the most parsimonious tree for complete sequences (Supplementary Figure 4), within sub-haplogroup N1b1a2, a transition at position m.204T>C defines a cluster with three samples, one from this study along with one from Zamora and another with an unknown origin. To further investigate this cluster, a search for similar CR haplotypes was performed in the EMPOP database and only three sequences were found. Two of them were from the same geographical area, namely Zamora (the same sample from Supplementary Figures 3 and 4) and Miranda do Douro,41, 42 both places with a well-documented history of Jewish presence. The remaining one was interestingly also from a Jew, an Ashkenazi from Hungary,62 thus this transition could be identified as a Jewish, albeit not exclusively Sephardic founding lineage.
In the N1b1a5 sub-clade, the only matches found at EMPOP database, including the transition at position m.16311T>C, were with five samples from Miranda do Douro41 and one from Uzbekistan,63 about which no further information was available. As neither the Bragança Jews nor the Mirandese population share haplotypes with the Portuguese population, and given their geographic proximity in a rather remote and isolated area, as well as the fact that there was an organized Jewish community in Miranda, at least from the twelfth century on,4 these individuals could easily also be Jewish descendants who lost memory of their origins or have not been detected as such, as in Mairal et al41 sampling did not include ethnical criteria.
Regarding the N1a1a1a2 branch, the transitions at positions m.150C>T, m.4501C>T and m.11977C>T defines a cluster that includes the Bragança Jews (with no shared haplotypes with the Portuguese population). Lineages carrying the m.150C>T transition were not found in available databases and thus represent what could be a specific feature of the Sephardic Jews from NE Portugal.
Considering that haplogroup N1 is very rare in contemporary European populations, the low number of complete available sequences from the Near East, as well as the poor definition obtained so far for this haplogroup in the Iberian Neolithic samples60 (N*), it seems at the moment hazardous to conclude whether the lineages found here are relics brought from the Near East by the first Jewish diasporas or were assimilated into the ancestral Iberian Jewish population in Europe
The T haplogroup in the Jewish population of Bragança is much more frequent than in Portugal40 and pertains to the T2 branch, with two different sub-clades: T2b11 and T2e1. It has been proposed that haplogroup T, which diverged from the macro haplogroup JT around 40 kya, underwent a dramatic expansion from its geographical origin in the Near East into Europe before the Neolithic period.64 An European indigenous dispersion has been proposed for T2 sub-clades, namely the T2b and T2e paraphyletic lineages.64 Recently, several mitochondrial lineages, defined as the ‘Neolithic package’ including T2, were also proposed to explain a rapid change from hunter-gatherering to farming, in the Early/Middle Neolithic, indicating a genetic flow from the Near East, Anatolia, and the Caucasus into Europe.65 T2b is the most common lineage among T2, reaching higher frequencies in Western Europe than in the Near East.64 Considering T2b branch, no haplotypes were shared between Jews and Portuguese individuals for the CR, demonstrating the absence of reciprocal introgression of T2b lineages between these two populations (Supplementary Figure 5).
Regarding the complete mtDNA genome, the Bragança samples nest in the branch defined by the transition at position m.9181A>G, along with a sample from Spain (Supplementary Figure 6). Interestingly, the Spanish sample is from the Zamora province (the same sample from Supplementary Figure 5) which, as stated above, is a geographically neighboring region to Bragança, at the Portuguese-Spanish border, and has also a well-documented Jewish past, with a well-established community dating back to 1259 CE.66 Hence, the m.9181A>G variant within the T2b11 branch seems to be a regionally specific variant within the T2b11 branch, reflecting a Sephardic signature, given its absence from public databases. The additional motif m.4902A>G-m.8557G>A-m.16167C>T-m.16261C>T is only present among the Portuguese Jews.
Haplogroup T2e is spread throughout southern Europe and the Mediterranean, but is also found in Scandinavia, Egypt, the Near East and Arabia64 and has been described as a founding lineage in the Bulgarian Sephardic community.12 More recently, it was also found among Sephardic descendants in Turkey, in Northern Mexico and south Texas, USA, being interpreted as a ‘Sephardic signature’ inside haplogroup T.31, 32 According to the more recent nomenclature in PhyloTree built 16,37 T2e’s sub-branches, T2e1 and T2e1a, are defined by the variants m.41C>T and m.2308A>G, respectively. Considering the growing number of complete mtDNA sequences available at this time, it was possible to define a new sub-haplogroup T2e1a1, based on m.15499C>T variant. It is important to notice that variants at position m.41C>T are exceptionally infrequent along the mitochondrial phylogeny. Position m.41C>T seems quite unstable inside the new defined T2e1a1 sub-branch (Supplementary Figure 6). Considering a back-mutation at position m.41C>T and taking all mtDNA genomes available to date, the m.2308A>G-m.15499C>T-m.16114C>T-m.16192C>T motif defines one of the branches (T2e1a1a1) of Portuguese Sephardic signature within T2e, previously reported for a Sephardic sample32 from Turkey, one of the Mediterranean countries that received exiled Iberian Jews. Two other samples shared this same motif, one from Mexico and another one from Texas. Without m.16114C>T, there was also a sample from Mexico. All these samples are from the study of Bedford et al31 and although the ethnicity of these three samples is not known, their Iberian ancestry seems consistent with a Sephardic origin. The Bragança Jews present two further distinct variants in the Sephardic signature, m.13135G>A and m.7133C>T, the latter not described until now. In conclusion, the back-mutation at position 41 inside this new sub-haplogroup T2e1a1 is entirely associated, so far, with Sephardic or probable Sephardic ancestry.
Inside the T2e1 branch, a new sub-haplogroup T2e1b (Supplementary Figure 6), defined by the presence of variant m.9181A>G, has been proposed in PhyloTree built 16.37 T2e1b is supported by 11 complete mtDNA sequences, including the Bragança Jews. Except for two samples from mtDNA Community database,67 without information concerning their ethnicity, all the remaining nine individuals are Jews, Sephardim or Ashkenazim.
The last putative Jewish Sephardic founding lineage belongs to haplogroup U2, particularly its European U2e sub-haplogroup. This lineage is extremely rare in modern European populations and was recently found in Late Neolithic68 and Iron Age populations, especially in the north of Europe.69 It also appears with rather high frequencies in Croatia,70 western Eurasia71 and the Basque country.50, 72 All U2e individuals in this study belong to the U2e1a1 sub-clade. A high frequency of haplogroup U2 was found in the Bragança Jews (7.0%), whereas in Portugal it only reaches a frequency of 1.3%, being found absent in the Bragança district.40 U2e1 lineages found in the Bragança Jewish population are shared with two individuals from Miranda; accordingly, this is the only putative Jewish founding lineage shared with the Portuguese population, when only the CR is looked upon (Supplementary Figure 7).
However, when considering complete mtDNA sequences, the Bragança clade is isolated from all other U2e1a1 haplotypes, except for another Jewish Ashkenazi sample from Moldova (Supplementary Figure 8), sharing the motif m.8014A>G—m.13708G>A, seemingly Jewish specific.
The strong founder effect previously reported in the maternal lineages of Portuguese Jews from Belmonte12 cannot be considered a general trait of the Sephardic groups in Portugal. In fact, NE communities, despite preserving a distinctive lineage profile, displayed diversity levels similar to the host population. Remarkably, for two of the founder lineages (T2e1b and U2e1a), defined by the complete mitochondrial genome, the shared sequences belong to both Sephardic as well as Ashkenazi Jews. Two possible scenarios could accommodate this finding: either the defining variants for each branch could have arisen before the separation between the two Jewish groups; or there may have been recent introgression of Sephardic lineages into Ashkenazim communities in the north of Europe. More complete sampling and complete sequences will contribute in the clarification of which one is more likely. In any case, it must be said that although not frequent, marriages between the two communities occurred (especially) in the sixteenth and seventeenth centuries, namely among the elite sugar traders, with the descendants assimilated into the Ashkenazi community.73, 74
The estimates of both the diversity levels and the number of female effective-population founders point at a stable size of the studied populations, in agreement with previous findings for the male counterparts.27 As expected, it was possible to identify some Sephardic signatures as well as signs of introgression from the host non-Jewish population. This gene flow seems to have been mutual as the maintenance of the observed diversity levels can only be explained by a number of founders that is higher than the Jewish families, which can still be traced today.
A study of recombinant markers is now required to unveil the reproductive strategies that have sustained this ancestral signature along with a moderate degree of admixture with the host population.
Accession codes
References
Tavares MJPF : A herança judaica em Portugal. Lisboa: CTT Correios de Portugal, 2004.
Martins J : Portugal e os judeus: Judaísmo e anti-semitismo no século XX. Lisboa: Vega, 2006.
Garcia MA : Os judeus de Belmonte: os caminhos da memória. Lisboa: Instituto de Sociologia e Etnologia das Religiões, Universidade Nova de Lisboa, 1993.
Pignatelli M : A comunidade israelita de Lisboa: o passado e o presente na construcção da etnicidade dos judeus de Lisboa. Lisboa: Universidade Técnica de Lisboa, Instituto Superior de Ciências Sociais e Políticas, 2000.
Saraiva AJ, Salomon HP, Sassoon ISD The Marrano Factory: The Portuguese Inquisition and Its New Christians 1536–1765. Leiden: Brill 2001.
Azevedo EM Cristãos Novos No Nordeste Transmontano: Los caminos de Cervantes y Sefarad: actas del II Congreso Internacional. Zamora, Bragança, Puebla de Sanabria, Benavente: Asociación Caminos de Cervantes y Sefarad 1995.
Mea E O Judaísmo no Séc. XX—A fénix renascida em Trás-os-Montes: Congresso Histórico 450 Anos da Fundação da Diocese de Bragança-Miranda. Bragança: Fundação da Diocese de Bragança-Miranda 1996.
Mea EA : A inquisição de Coimbra no século XVI: a instituição, os homens e a sociedade. Porto: Fundação Engo António de Almeida, 1997.
Paulo A : Os judeus secretos em Portugal. Fafe: Editorial Labirinto, 1985.
Adams SM, Bosch E, Balaresque PL et al: The genetic legacy of religious diversity and intolerance: paternal lineages of Christians, Jews, and Muslims in the Iberian Peninsula. Am J Hum Genet 2008; 83: 725–736.
Bauchet M, McEvoy B, Pearson LN et al: Measuring European population stratification with microarray genotype data. Am J Hum Genet 2007; 80: 948–956.
Behar DM, Metspalu E, Kivisild T et al: Counting the founders: the matrilineal genetic ancestry of the Jewish Diaspora. PLoS One 2008; 3: e2062.
Hammer MF, Redd AJ, Wood ET et al: Jewish and Middle Eastern non-Jewish populations share a common pool of Y-chromosome biallelic haplotypes. Proc Natl Acad Sci USA 2000; 97: 6769–6774.
Kopelman NM, Stone L, Wang C et al: Genomic microsatellites identify shared Jewish ancestry intermediate between Middle Eastern and European populations. BMC Genet 2009; 10: 80.
Olshen AB, Gold B, Lohmueller KE et al: Analysis of genetic variation in Ashkenazi Jews by high density SNP genotyping. BMC Genet 2008; 9: 14.
Ostrer H : A genetic profile of contemporary Jewish populations. Nat Rev 2001; 2: 891–898.
Price AL, Butler J, Patterson N et al: Discerning the ancestry of European Americans in genetic association studies. PLoS Genet 2008; 4: e236.
Seldin MF, Shigeta R, Villoslada P et al: European population substructure: clustering of northern and southern populations. PLoS Genet 2006; 2: e143.
Campbell CL, Palamara PF, Dubrovsky M et al: North African Jewish and non-Jewish populations form distinctive, orthogonal clusters. Proc Natl Acad Sci USA 2012; 109: 13865–13870.
Ostrer H, Skorecki K : The population genetics of the Jewish people. Hum Genet 2013; 132: 119–127.
Atzmon G, Hao L, Pe’er I et al: Abraham’s children in the genome era: major Jewish diaspora populations comprise distinct genetic clusters with shared Middle Eastern Ancestry. Am J Hum Genet 2010; 86: 850–859.
Behar DM, Yunusbayev B, Metspalu M et al: The genome-wide structure of the Jewish people. Nature 2010; 466: 238–242.
Velez C, Palamara PF, Guevara-Aguirre J et al: The impact of Converso Jews on the genomes of modern Latin Americans. Hum Genet 2012; 131: 251–263.
Gerber S, Rozet JM, Takezawa SI et al: The photoreceptor cell-specific nuclear receptor gene (PNR) accounts for retinitis pigmentosa in the Crypto-Jews from Portugal (Marranos), survivors from the Spanish Inquisition. Hum Genet 2000; 107: 276–284.
Sutton WK, Knight A, Underhill PA, Neulander JS, Disotell TR, Mountain JL : Toward resolution of the debate regarding purported crypto-Jews in a Spanish-American population: evidence from the Y chromosome. Ann Hum Biol 2006; 33: 100–111.
Bordenave K, Griffith J, Hordes SM, Williams TM, Padilla RS : The historical and geomedical immunogenetics of pemphigus among the descendants of Sephardic Jews in New Mexico. Arch Dermatol 2001; 137: 825–826.
Nogueiro I, Manco L, Gomes V, Amorim A, Gusmao L : Phylogeographic analysis of paternal lineages in NE Portuguese Jewish communities. Am J Phys Anthropol 2010; 141: 373–381.
Picornell A, Gomez-Barbeito L, Tomas C, Castro JA, Ramon MM, Mitochondrial DNA : HVRI variation in Balearic populations. Am J Phys Anthropol 2005; 128: 119–130.
Tomas C, Picornell A, Castro JA et al: Genetic variability at nine STR loci in the Chueta (Majorcan Jews) and the Balearic populations investigated by a single multiplex reaction. Int J Legal Med 2000; 113: 263–267.
Tomas C, Picornell A, Castro JA, Ramon MM : Genetic analyses in five western Mediterranean populations: variation at five tetrameric short tandem repeat loci. Hum Biol 2001; 73: 349–363.
Bedford FL, Yacobi D, Felix G, Garza FM : Clarifying Mitochondrial DNA Subclades of T2e from Mideast to Mexico. J Phylogenetics Evol Biol 2013; 1: 121.
Bedford FL : Sephardic signature in haplogroup T mitochondrial DNA. Eur J Hum Genet 2012; 20: 441–448.
Teixeira JC, Nogueiro I, Goios A, Gusmão L, Amorim A, Alvarez L : Mitochondrial DNA-control region sequence variation in the NE Portuguese Jewish community. FSI Genet Suppl Ser 2011; 3: e51–e52.
Ramos A, Santos C, Alvarez L, Nogues R, Aluja MP : Human mitochondrial DNA complete amplification and sequencing: a new validated primer set that prevents nuclear DNA sequences of mitochondrial origin co-amplification. Electrophoresis 2009; 30: 1587–1593.
Ramos A, Santos C, Barbena E et al: Validated primer set that prevents nuclear DNA sequences of mitochondrial origin co-amplification: a revision based on the New Human Genome Reference Sequence (GRCh37). Electrophoresis 2011; 32: 782–783.
Andrews RM, Kubacka I, Chinnery PF, Lightowlers RN, Turnbull DM, Howell N : Reanalysis and revision of the Cambridge reference sequence for human mitochondrial DNA. Nat Genet 1999; 23: 147.
van Oven M, Kayser M : Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation. Hum Mutat 2009; 30: E386–E394.
Parson W, Dur A : EMPOP—a forensic mtDNA database. Forensic Sci Int Genet 2007; 1: 88–92.
Excoffier L, Lischer HE : Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol Ecol Resour 2010; 10: 564–567.
Pereira L, Prata MJ, Amorim A : Diversity of mtDNA lineages in Portugal: not a genetic edge of European variation. Ann Hum Genet 2000; 64: 491–506.
Mairal Q, Santos C, Silva M et al: Linguistic isolates in Portugal: insights from the mitochondrial DNA pattern. Forensic Sci Int Genet 2013; 7: 618–623.
Ramos A, Santos C, Mateiu L et al: Frequency and pattern of heteroplasmy in the complete human mitochondrial genome. PLoS One 2013; 8: e74636.
IBM Corp. Released: IBM SPSS Statistics for Windows, Version 21.0. Armonk, NY: IBM Corp,, 2012.
Bandelt HJ, Forster P, Sykes BC, Richards MB : Mitochondrial portraits of human populations using median networks. Genetics 1995; 141: 743–753.
Zhang C, Huang VH, Simon M et al: Heteroplasmic mutations of the mitochondrial genome cause paradoxical effects on mitochondrial functions. FASEB J 2012; 26: 4914–4924.
Helgason A, Sigureth ardottir S, Gulcher JR, Ward R, Stefansson K : mtDNA and the origin of the Icelanders: deciphering signals of recent population history. Am J Hum Genet 2000; 66: 999–1016.
Pereira L, Cunha C, Amorim A : Predicting sampling saturation of mtDNA haplotypes: an application to an enlarged Portuguese database. Int J Legal Med 2004; 118: 132–136.
Torroni A, Bandelt HJ, Macaulay V et al: A signal, from human mtDNA, of postglacial recolonization in Europe. Am J Hum Genet 2001; 69: 844–852.
Garcia O, Fregel R, Larruga JM et al: Using mitochondrial DNA to test the hypothesis of a European post-glacial human recolonization from the Franco-Cantabrian refuge. Heredity (Edinb) 2011; 106: 37–45.
Cardoso S, Alfonso-Sanchez MA, Valverde L et al: The maternal legacy of Basques in northern navarre: New insights into the mitochondrial DNA diversity of the Franco-Cantabrian area. Am J Phys Anthropol 2011; 145: 480–488.
Alvarez L, Santos C, Ramos A et al: Mitochondrial DNA patterns in the Iberian Northern plateau: Population dynamics and substructure of the Zamora province. Am J Phys Anthropol 2010; 142: 531–539.
Costa MD, Pereira JB, Pala M et al: A substantial prehistoric European ancestry amongst Ashkenazi maternal lineages. Nat Commun 2013; 4: 2543.
Macaulay V, Hill C, Achilli A et al: Single, rapid coastal settlement of Asia revealed by analysis of complete mitochondrial genomes. Science 2005; 308: 1034–1036.
Fernandes V, Alshamali F, Alves M et al: The Arabian cradle: mitochondrial relicts of the first steps along the southern route out of Africa. Am J Hum Genet 2012; 90: 347–355.
Haak W, Balanovsky O, Sanchez JJ et al: Ancient DNA from European early neolithic farmers reveals their near eastern affinities. PLoS Biol 2010; 8: e1000536.
Palanichamy MG, Zhang CL, Mitra B et al: Mitochondrial haplogroup N1a phylogeography, with implication to the origin of European farmers. BMC Evol Biol 2010; 10: 304.
Deguilloux MF, Soler L, Pemonge MH, Scarre C, Joussaume R, Laporte L : News from the west: ancient DNA from a French megalithic burial chamber. Am J Phys Anthropol 2011; 144: 108–118.
Sampietro ML, Lao O, Caramelli D et al: Palaeogenetic evidence supports a dual model of Neolithic spreading into Europe. Proc R Soc B 2007; 274: 2161–2167.
Hervella M, Izagirre N, Alonso S et al: Ancient DNA from hunter-gatherer and farmer groups from Northern Spain supports a random dispersion model for the Neolithic expansion into Europe. PLoS One 2012; 7: e34417.
Gamba C, Fernandez E, Tirado M et al: Ancient DNA from an Early Neolithic Iberian population supports a pioneer colonization by first farmers. Mol Ecol 2012; 21: 45–56.
Behar DM, Metspalu E, Kivisild T et al: The matrilineal ancestry of Ashkenazi Jewry: portrait of a recent founder event. Am J Hum Genet 2006; 78: 487–497.
Brandstatter A, Egyed B, Zimmermann B, Tordai A, Padar Z, Parson W : Mitochondrial DNA control region variation in Ashkenazi Jews from Hungary. Forensic Sci Int Genet 2008; 2: e4–e6.
Irwin JA, Ikramov A, Saunier J et al: The mtDNA composition of Uzbekistan: a microcosm of Central Asian patterns. Int J Legal Med 2010; 124: 195–204.
Pala M, Olivieri A, Achilli A et al: Mitochondrial DNA signals of late glacial recolonization of Europe from near eastern refugia. Am J Hum Genet 2012; 90: 915–924.
Brandt G, Haak W, Adler CJ et al: Ancient DNA reveals key stages in the formation of central European mitochondrial genetic diversity. Science 2013; 342: 257–261.
Casar MFG : El pasado judío de Zamora. Valladolid: Junta de Castilla y León, Consejería de Cultura y Turismo, 1992.
Behar DM, van Oven M, Rosset S et al: A ‘copernican’ reassessment of the human mitochondrial DNA tree from its root. Am J Hum Genet 2012; 90: 675–684.
Lee EJ, Makarewicz C, Renneberg R et al: Emerging genetic patterns of the european neolithic: perspectives from a late neolithic bell beaker burial site in Germany. Am J Phys Anthropol 2012; 148: 571–579.
Melchior L, Gilbert MT, Kivisild T, Lynnerup N, Dissing J : Rare mtDNA haplogroups and genetic differences in rich and poor Danish Iron-Age villages. Am J Phys Anthropol 2008; 135: 206–215.
Jeran N, Havas Augustin D, Grahovac B et al: Mitochondrial DNA heritage of Cres Islanders—example of Croatian genetic outliers. Coll Antropol 2009; 33: 1323–1328.
Quintana-Murci L, Chaix R, Wells RS et al: Where west meets east: the complex mtDNA landscape of the southwest and Central Asian corridor. Am J Hum Genet 2004; 74: 827–845.
Cardoso S, Villanueva-Millan MJ, Valverde L et al: Mitochondrial DNA control region variation in an autochthonous Basque population sample from the Basque Country. Forensic Sci Int Genet 2012; 6: e106–e108.
Roth C, Spivak A Historia de los marranos. Editorial. Israel 1946.
Mea E, Problemática do Judaísmo. A Séculos XVI-XVII: Congresso Internacional Inquisição Portuguesa. Tempo, Razão e Circunstância. Lisboa, S Paulo, 2007.
Acknowledgements
We thank Walther Parson and Mannis van Oven for the helpful discussion and suggestions in the phylogenetic analysis, Doron Behar and Felice L Bedford for the additional information concerning Jewish samples. Elvira Mea, Jorge Martins, Fernanda Guimarães, Jits van Straten and Harmen Snel for their contribution on Jewish history. Kevin Charles Rowe for the English language revision and Sofia Marques for the aid in the preparation of figures. All DNA donors for their collaboration and generosity. This work was supported by FCT, Portuguese Foundation for Science and Technology, financed by the European Social Funds (COMPETE-FEDER), project PTDC/ATP-DEM/4545/2012. LA (SFRH/BPD/65000/2009), IN (SFRH/BD/73336/2010) and JT (SFRH/BD/77043/2011) are supported by FCT fellowships and funded by POPH-QREN—Promotion of scientific employment, supported by the European Social Fund and national funds of the Ministry of Education and Science. IPATIMUP is an Associate Laboratory of the Portuguese Ministry of Science, Technology and Higher Education and is partially supported by FCT.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on European Journal of Human Genetics website
Rights and permissions
About this article

Cite this article
Nogueiro, I., Teixeira, J., Amorim, A. et al. Echoes from Sepharad: signatures on the maternal gene pool of crypto-Jewish descendants. Eur J Hum Genet 23, 693–699 (2015). https://doi.org/10.1038/ejhg.2014.140
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ejhg.2014.140
This article is cited by
-
Middle eastern genetic legacy in the paternal and maternal gene pools of Chuetas
Scientific Reports (2020)
-
On two Jewish clades in mitochondrial DNA
European Journal of Human Genetics (2015)
-
Reply to letter from Felice L. Bedford and Doron Yacobi
European Journal of Human Genetics (2015)
