
Abstract
We used exome sequencing to study a non-consanguineous family with two children who had anterior segment dysgenesis, sclerocornea, microphthalmia, hypotonia and developmental delays. Sanger sequencing verified two Peroxidasin (PXDN) mutations in both sibs—a maternally inherited, nonsense mutation, c.1021C>T predicting p.(Arg341*), and a paternally inherited, 23-basepair deletion causing a frameshift and premature protein truncation, c.2375_2397del23, predicting p.(Leu792Hisfs*67). We re-examined exome data from 20 other patients with structural eye defects and identified two additional PXDN mutations in a sporadic male with bilateral microphthalmia, cataracts and anterior segment dysgenesis—a maternally inherited, frameshift mutation, c.1192delT, predicting p.(Tyr398Thrfs*40) and a paternally inherited, missense substitution that was predicted to be deleterious, c.947 A>C, predicting p.(Gln316Pro). Mutations in PXDN were previously reported in three families with congenital cataracts, microcornea, sclerocornea and developmental glaucoma. The gene is expressed in corneal epithelium and is secreted into the extracellular matrix. Defective peroxidasin has been shown to impair sulfilimine bond formation in collagen IV, a constituent of the basement membrane, implying that the eye defects result because of loss of basement membrane integrity in the developing eye. Our finding of a broader phenotype than previously appreciated for PXDN mutations is typical for exome-sequencing studies, which have proven to be highly effective for mutation detection in patients with atypical presentations. We conclude that PXDN sequencing should be considered in microphthalmia with anterior segment dysgenesis.
Similar content being viewed by others


INTRODUCTION
Anophthalmia (absent eyes), microphthalmia (small eyes) and coloboma (defective closure of the optic fissure) are important structural birth defects because of the medical significance of severely reduced vision. Mutations in several genes, including SOX2, ALDH1A3, STRA6, OTX2, RAX, BMP4, VSX2, GDF6, SIX6, PAX6, VAX1 and SMOC1 can cause anophthalmia and microphthalmia (A/M), but there is substantial genetic heterogeneity and >50% of affected individuals do not receive a molecular genetic diagnosis for their birth defect after testing of the currently known causative genes.1, 2, 3, 4, 5, 6, 7, 8 A/M can occur as an isolated malformation (simplex A/M) or can be combined with additional ocular abnormalities affecting anterior or posterior segments of the eye (complex A/M). Complex A/M often includes anterior segment dysgenesis, an eye defect constituting maldevelopment of the anterior structures behind the cornea and in front of the lens, including the iris, ciliary body and trabecular meshwork. Anterior segment dysgenesis is important to recognize because it is frequently associated with cataracts and predisposes to increased ocular pressure and developmental glaucoma in 50% of cases.9 FOXC1 and PITX2 disruptions, either mutations or deletions, have been estimated to account for 6–40% of patients with anterior segment dysgenesis,10, 11 but are frequently associated with systemic findings, such as dental and umbilical anomalies in patients with PITX2 haploinsufficiency and cardiac and hearing deficits in patients with FOXC1 haploinsufficiency.11 Mutations in FOXE3, PITX3, PAX6, B3GALTL, MAF, COL4A1, CYP1B1 and SH3PXD2B have also been described in patients with anterior segment dysgenesis,9, 12, 13, 14, 15, 16, 17, 18, 19 but there is considerable genetic heterogeneity similar to A/M and many genes remain unidentified. Next-generation sequencing has proven to be an outstandingly successful technology for new gene identification,20 and we present our results for two families, a sibship with complex microphthalmia and anterior segment dysgenesis and a sporadic male patient with unilateral microphthalmia anterior segment dysgenesis. In both families, exome sequencing identified PXDN mutations as a cause for their eye defects.
MATERIALS AND METHODS
Clinical reports
In the first family, the older male sibling (family 1, patient (P); Figure 1a) had bilateral microphthalmia and sclerocornea with iridocorneal dysgenesis and glaucoma. The right eye was more severely affected than the left eye. He had persistent hypotonia that was first diagnosed in the neonatal period and had experienced mild gross motor and fine motor delays. He rolled over at 5 months, sat at 9 months and walked at 26 months of age. He required occupational therapy for his motor delays. He spoke in phrases at 12 months of age and was educated in a mainstream classroom with support for visual impairment. His younger sister (family 1, sibling (S); Figure 1b) had bilateral microphthalmia, sclerocornea and anterior segment dysgenesis. She rolled over at 6 months and cruised at 18 months of age. She did not have phrases at 2 years of age and received speech therapy. She has had hypotonia and required occupational therapy for delays in fine motor skills and tactile defensiveness. An evaluation by early intervention services at 24 months of age determined her development to be compatible with 12 months chronological age. There was no other history of eye defects in the family and no known consanguinity. Ethnicity was Caucasian.

(a) Frontal view of the male sibling with PXDN mutations c.1021C>T predicting p.(Arg341*) and c.2375_2397del23, predicting p.(Leu792Hisfs*67), showing bilateral scleral opacities. (b) Frontal view of the female sibling with PXDN mutations c.1021C>T predicting p.Arg341* and c.2375_2397del23, predicting p.(Leu792Hisfs*67), showing bilateral scleral opacities and right microphthalmia.
The second patient was a Caucasian male with bilateral anterior segment dysgenesis and left microphthalmia diagnosed in the newborn period. He had a moderate sized cataract in his left eye and a small cataract in his right eye and underwent left lensectomy and pupilloplasty at 11 days of age. His development was normal at 3 years and 4 months of age and a magnetic resonance imaging scan of the brain was reported as unremarkable. His maternal half brother had amblyopia and a small nodule on the helix of his right ear. Both parents were healthy with no history of eye defects. PITX2 and FOXC1 sequencing returned normal, and array comparative genomic hybridization (180K, Agilent, Santa Clara, CA, USA) as a clinical test was unrevealing. There was no known consanguinity.
Exome sequencing
Ethical approval for the study was obtained from the Committee on Human Research at the University of California, San Francisco. After obtaining written, informed consent, venous bloods were obtained and DNA was extracted from all three affected individuals and both sets of parents. Exome sequencing was performed as previously described.7, 21 Briefly, libraries were prepared using biotinylated DNA oligonucleotides (SeqCap EZ Human Exome Library v3.0; Roche Nimblegen, Madison, WI, USA) and sequencing was performed on a HiSeq2000 (Illumina, San Diego, CA, USA) for paired-end 100 cycles. Sequencing reads were aligned to the hg19 reference genome using the Burrows–Wheeler Alignment tool (BWAv0.5.9) using the default parameters that allow two mismatches. Indexing, realignment and duplicate removal were performed using Picard and Samtools, and variants were subsequently called using the Genome Analysis Toolkit v 1.3-21-gcb284ee. We then utilized wAnnovar (http://wannovar.usc.edu/) with default parameters to analyze the sequence variants.22, 23 We favored autosomal recessive inheritance because the first family had an affected male and female sibling pair, although we considered all possible modes of inheritance to analyze the data. Novel sequence variants were assessed for deleterious consequences using Polyphen-2 (http://genetics.bwh.harvard.edu/pph2/) and Mutation Taster (http://mutationtaster.org/MutationTaster/).24 Selected novel sequence variants were verified using Sanger sequencing. After verification of the two PXDN mutations in the first sibship, we re-examined our existing exome sequencing data from 20 patients with developmental eye defects for PXDN sequence alterations, and detected two predicted mutations in the sporadic male case that were subsequently verified by Sanger sequencing. We also used Sanger sequencing to interrogate PXDN for mutations in 13 additional patients with anophthalmia or microphthalmia who were selected because of abnormal anterior segment findings, including sclerocornea, microcornea or cataract (Supplementary Table S1). These patients had not undergone exome sequencing. The 13 additional A/M cases that we sequenced were ascertained by the population-based surveillance program—California Birth Defects Monitoring Program.25 These A/M cases were linked to their newborn bloodspots, which represented the source of DNA. All samples were obtained with the approval form the State of California Health and Welfare Agency for the Protection of Human Subjects.
RESULTS
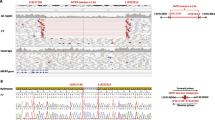
Coverage for each of the exomes is shown in Supplementary Table S2 and the filtering strategy is summarized in Supplementary Table S3. In the oldest sib from the first family, exome sequencing revealed 21 616 sequence variants, whereas the second sib had 21 995 sequence variants. After removing sequence variants in the database of single-nucleotide polymorphisms (http://www.ncbi.nlm.nih.gov/projects/SNP/) and variants in unknown genes, variants in 262 genes were shared by both sibs. Of these genes, 72 had synonymous variants and were not further considered. Genes (138) with only one shared variant were also not further considered. Of the remaining genes, 11 had exactly two novel sequence variants and four of these 11 genes were expressed in the eye (NEIBank; http://neibank.nei.nih.gov/index.shtml; data not shown). From the four genes, only PXDN was known to be involved in developmental eye defects.26 The predicted mutations in PXDN (mRNA reference sequence NM_012293.1, Chromosome reference number NC_000002.11, National Center for Biotechnology Information Gene, http://www.ncbi.nlm.nih.gov/gene; Ensembl genomic reference ENSG00000130508 and transcript ENST00000252804; http://www.ensembl.org/index.html) in the first family were verified by Sanger sequencing: a maternally inherited, nonsense mutation: c.1021C>T, predicting p.(Arg341*) and a paternally inherited, intragenic deletion resulting in a frameshift and premature protein truncation, c.2375_2397del23, predicting p.(Leu792Hisfs*67) (Figures 2a,b and Figure 3a). Re-examination of our existing exome data from 20 other families identified two additional PXDN mutations in a sporadic male patient, which were also confirmed with Sanger sequencing—a paternally inherited, missense mutation, c.947A>C, predicting p.(Gln316Pro), together with a maternally inherited, frameshift mutation, c.1192delT, predicting p.(Tyr398Thrfs*40) (Figures 2c,d and Figure 3a). The missense mutation p.(Gln316Pro) was predicted to be probably damaging with Polyphen-2 (score 0.961; sensitivity 0.78 with specificity 0.95) and to be disease causing by Mutation Taster (P=0.999). It is likely that the pathogenic effect of the mutation results from the substitution of the amino acid, as the glutamine residue was conserved in chimp, mouse, chicken and cat (Figure 3b). Neither mutation was present in the Exome Variant Server database (http://evs.gs.washington.edu/EVS/). Of the four mutations that we identified, three were novel, whereas p.(Arg341*) has been previousIy published.26 In the other individuals with A/M selected because of additional anterior segment abnormalities, we did not find two pathogenic sequence alterations in any patient. Mutations have been submitted to the Leiden Open Variation Database (http://databases.lovd.nl/shared/genes/PXDN).

(a) Chromatograms showing c.1021C>T predicting p.(Arg341*) in PXDN in the patient (P) of the first family, sister of the patient (S) and mother of the patient (M). The father of the patient (F) does not have the mutation. (b) Chromatograms showing the start of mutation c.2375_2397del23, predicting p.(Leu792Hisfs*67) in PXDN in the patient (P), sister of the patient (S) and father of the patient (F). The mother of the patient (M) does not have the mutation. (c) Chromatograms showing c.947A>C, predicting p.(Gln316Pro) in PXDN in the patient (P) of the second family and the father of the patient (F). The mother of the patient (M) does not have the mutation. (d) Chromatograms showing c.1192delT, predicting p.(Tyr398Thrfs*40) in PXDN in the patient (P) of the second family and the mother of the patient (M). The father of the patient (F) does not have the mutation.

(a) Diagram of peroxidasin showing domain location of PXDN mutations described to date. Adapted from reference;26 not drawn to scale. Figures above domains represent amino-acid numbers. The mutations in bold were described in this paper and the mutations in italics were described in Khan et al.26 (b) Conservation of amino-acid residues at p.Gln(Q)316, the site of c.947A>C and p.(Gln316Pro). The glutamine residue is conserved in all species with this residue.
DISCUSSION
Mutations causing PXDN loss of function have previously been described in three families who had congenital cataract-microcornea with corneal opacity (CCMCO; Table 1).26 Defining phenotypic features for these families were microcornea and corneal opacification, with varying degrees of intrafamilial expression, but not microphthalmia. A fourth family with PXDN loss of function had developmental glaucoma, buphthalmos and extensive corneal opacification.26 No patient had neurological abnormalities as seen in our first family. In all reported families, unaffected carriers have had no ocular abnormalities.
The human PXDN gene has 23 exons and encodes a 1479-amino-acid protein. At embryonic day (E) 18.5, Pxdn is expressed in the corneal endothelium and in adult murine eyes, and Pxdn is present in the corneal and lens epithelium.26 PXDN was weakly expressed in normal brain tissue and was upregulated in the microvasculature and glioma endothelial cells of primary glial and metastatic brain tumors.27 Expression has also been demonstrated in several other tumor types including melanoma, breast, ovarian, colon and renal cell cancer.28, 29 PXDN encodes peroxidasin, a protein that is located in the endoplasmic reticulum and secreted into the extracellular space. It contains a peroxidase domain common to other mammalian peroxidases, for example, myeloperoxidase and thyroid peroxidase, and other motifs typical for extracellular proteins, including an amino-terminal secretory signal sequence and ligand-binding domains comprising leucine-rich repeat regions, C2-type immunoglobulin-like motifs and a von Willebrand factor type-C domain.26 In the mutations identified to date, p.(Gln316Pro) affects a residue in an immunoglobulin-like domain, and this glutamine residue is highly conserved (Figure 3). The stop mutation p.(Arg341*) and frameshift mutation p.(Tyr398Thrfs*40) are not present in named protein domains, but both mutations are predicted to cause nonsense-mediated decay. The remaining mutations, p.(Leu792Hisfs*67), p.(Cys857Alafs*5) and p.(Arg880Cys), occur at residues in the peroxidase domain, and the first two mutations predict nonsense-mediated decay (Figure 3a). At present, there is no phenotype–genotype correlation, and although the first sibship in this report had two truncating mutations, patients with homozygosity for other truncating mutations have not had similar additional findings of microphthalmia or developmental delays.26
Peroxidasins are involved in hydrogen peroxidase metabolism, and peroxidasin has been shown to catalyze the formation of a sulfilimine bond between apposed lysine and methionine residues in the C-terminal interface of collagen IV, the predominant constituent of the basement membrane.30 Loss of peroxidasin resulted in defective basement membrane integrity in Drosophila melanogaster and Caenorhabditis elegans mutants, and it is most likely that the PXDN mutations in human patients compromise the integrity of the basement membrane of the lens and cornea by disrupting collagen IV formation.30 Mutations resulting in haploinsufficiency for collagen IV have been shown to cause anterior segment defects in mice and in humans, including cataracts, corneal opacities, iris defects, iridocorneal adhesions and buphthalmos,31, 32, 33 similar to the eye malformations seen in patients with PXDN loss of function.
In the first family with two siblings, both were found to have developmental delays and hypotonia, although these manifestations have not been described in other reported patients and were not present in the second patient in this report. PXDN is weakly expressed in the brain and it is plausible that collagen IV and basement membrane formation could also be disrupted in the brain in patients with loss of PXDN function. Haploinsufficiency for collagen IV caused type I porencephaly and intracranial hemorrhage in human patients and cortical lamination defects in mice that can be associated with developmental differences.33, 34 However, it is still speculative as to whether the PXDN mutations are responsible for the hypotonia and delays in this family, although we were unable to find any other mutations to explain this aspect of their phenotype (data not shown). A duplication involving the entire PXDN gene was present in two male half-siblings with autism and inherited from their mother, who had mosaicism for the duplication.35 However, the duplication also disrupted MYT1L, a gene expressed in the brain and linked with schizophrenia and depression,36 and aberrant MYTL function was presumed to be responsible for the autism diagnosis in these children.
Our finding of a broader phenotype involving microphthalmia in patients with PXDN loss of function is characteristic for exome sequencing, which has proven to be highly effective for mutation detection in atypical presentations. We conclude that PXDN mutations should be considered in microphthalmia with anterior segment dysgenesis in addition to CCMCO.
References
Fantes J, Ragge NK, Lynch SA et al: Mutations in SOX2 cause anophthalmia. Nat Genet 2003; 33: 461–463.
Schneider A, Bardakjian T, Reis LM, Tyler RC, Semina EV : Novel SOX2 mutations and genotype phenotype correlation in anophthalmia and microphthalmia. Am J Med Genet A 2009; 149A: 2706–2715.
Verma AS, Fitzpatrick DR : Anophthalmia and microphthalmia. Orphanet J Rare Dis 2009; 2: 47.
Bardakjian T, Weiss A, Schneider AS : Anophthalmia/microphthalmia overview; in Pagon RA, Adam MP, Ardinger HH, Bird TD, Dolan CR, Fong CT, Smith RJH, Stephens K, (eds): GeneReviews® [Internet]., Seattle (WA): University of Washington, Seattle 1993–2014.
Slavotinek AM : Eye development genes and known syndromes. Mol Genet Metab 2011; 104: 448–456.
Slavotinek AM, Chao R, Vacik T et al: VAX1 mutation associated with microphthalmia, corpus callosum agenesis, and orofacial clefting: the first description of a VAX1 phenotype in humans. Hum Mutat 2011; 33: 364–368.
Yahyavi M, Abouzeid H, Gawdat G et al: ALDH1A3 loss of function causes bilateral anophthalmia/microphthalmia and hypoplasia of the optic nerve and optic chiasm. Hum Mol Genet 2013; 2: 3250–3258.
Aldahmesh MA, Khan AO, Hijazi H, Alkuraya FS : Homozygous truncation of SIX6 causes complex microphthalmia in humans. Clin Genet 2013; 84: 198–199.
Reis LM, Semina EV : Genetics of anterior segment dysgenesis disorders. Curr Opin Ophthalmol 2011; 22: 314–324.
D'haene B, Meire F, Claerhout I et al: Expanding the spectrum of FOXC1 and PITX2 mutations and copy number changes in patients with anterior segment malformations. Invest Ophthalmol Vis Sci 2011; 52: 324–333.
Reis LM, Tyler RC, Volkmann Kloss BA et al: PITX2 and FOXC1 spectrum of mutations in ocular syndromes. Eur J Hum Genet 2012; 20: 1224–1233.
Semina EV, Brownell I, Mintz-Hittner HA et al: Mutations in the human forkhead transcription factor FOXE3 associated with anterior segment ocular dysgenesis and cataracts. Hum Mol Genet 2001; 10: 231–236.
Ali M, Buentello-Volante B, McKibbin M et al: Homozygous FOXE3 mutations cause non-syndromic, bilateral, total sclerocornea, aphakia, microphthalmia and optic disc coloboma. Mol Vis 2010; 16: 1162–1168.
Doucette L, Green J, Fernandez B et al: A novel, non-stop mutation in FOXE3 causes an autosomal dominant form of variable anterior segment dysgenesis including Peters anomaly. Eur J Hum Genet 2011; 9: 293–299.
Semina EV, Ferrell RE, Mintz-Hittner HA et al: A novel homeobox gene PITX3 is mutated in families with autosomal-dominant cataracts and ASMD. Nat Genet 1998; 19: 167–170.
Jamieson RV, Perveen R, Kerr B et al: Domain disruption and mutation of the bZIP transcription factor, MAF, associated with cataract, ocular anterior segment dysgenesis and coloboma. Hum Mol Genet 2002; 11: 33–42.
Weng J, Luo J, Cheng X et al: Deletion of G protein-coupled receptor 48 leads to ocular anterior segment dysgenesis (ASD) through down-regulation of Pitx2. Proc Natl Acad Sci USA 2008; 105: 6081–6086.
Mao M, Solivan-Timpe F, Roos BR et al: Localization of SH3PXD2B in human eyes and detection of rare variants in patients with anterior segment diseases and glaucoma. Mol Vis 2012; 18: 705–713.
Alzuhairy S, Abu-Amero KK, Al-Shahwan S, Edward DP : A novel CYP1B1 mutation with congenital glaucoma and total aniridia. Ophthalmic Genet 2013; 3: (epub ahead of print).
Bamshad MJ, Ng SB, Bigham AW et al: Exome sequencing as a tool for Mendelian disease gene discovery. Nat Rev Genet 2011; 12: 745–755.
Slavotinek AM, Mehrotra P, Nazarenko I et al: Focal facial dermal dysplasia, type IV, is caused by mutations in CYP26C1. Hum Mol Genet 2013; 22: 696–703.
Wang K, Li M, Hakonarson H : ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res 2010; 38: e164.
Chang X, Wang K : wANNOVAR: annotating genetic variants for personal genomes via the web. J Med Genet 2012; 49: 433–436.
Schwarz JM, Rödelsperger C, Schuelke M, Seelow D : MutationTaster evaluates disease-causing potential of sequence alterations. Nat Methods 2010; 7: 575–576.
Croen LA, Shaw GM, Jensvold NG, Harris JA : Birth defects monitoring in California: a resource for epidemiological research. Paediatr Perinat Epidemiol 1991; 5: 423–427.
Khan K, Rudkin A, Parry DA et al: Homozygous mutations in PXDN cause congenital cataract, corneal opacity, and developmental glaucoma. Am J Hum Genet 2011; 89: 464–473.
Liu Y, Carson-Walter EB, Cooper A et al: Vascular gene expression patterns are conserved in primary and metastatic brain tumors. J Neurooncol 2010; 99: 13–24.
Mitchell MS, Kan-Mitchell J, Minev B et al: A novel melanoma gene (MG50) encoding the interleukin 1 receptor antagonist and six epitopes recognized by human cytolytic T lymphocytes. Cancer Res 2000; 60: 6448–6456.
Castronovo V, Waltregny D, Kischel P et al: A chemical proteomics approach for the identification of accessible antigens expressed in human kidney cancer. Mol Cell Proteomics 2006; 5: 2083–2091.
Bhave G, Cummings CF, Vanacore RM et al: Peroxidasin forms sulfilimine chemical bonds using hypohalous acids in tissue genesis. Nat Chem Biol 2012; 8: 784–790.
Van Agtmael T, Schlötzer-Schrehardt U, McKie L et al: Dominant mutations of Col4a1 result in basement membrane defects which lead to anterior segment dysgenesis and glomerulopathy. Hum Mol Genet 2005; 14: 3161–3168.
Gould DB, Marchant JK, Savinova OV et al: Col4a1 mutation causes endoplasmic reticulum stress and genetically modifiable ocular dysgenesis. Hum Mol Genet 2007; 16: 798–807.
Labelle-Dumais C, Dilworth DJ, Harrington EP et al: COL4A1 mutations cause ocular dysgenesis, neuronal localization defects, and myopathy in mice and Walker-Warburg syndrome in humans. PLoS Genet 2011; 7: e1002062.
Kuo DS, Labelle-Dumais C, Gould DB : COL4A1 and COL4A2 mutations and disease: insights into pathogenic mechanisms and potential therapeutic targets. Hum Mol Genet 2012; 21: R97–110.
Meyer KJ, Axelsen MS, Sheffield VC et al: Germline mosaic transmission of a novel duplication of PXDN and MYT1L to two male half-siblings with autism. Psychiatr Genet 2012; 22: 137–140.
Lee Y, Mattai A, Long R et al: Microduplications disrupting the MYT1L gene (2p25.3) are associated with schizophrenia. Psychiatr Genet 2012; 22: 206–209.
Acknowledgements
We are grateful to the families for their participation. This work was supported by grant R21EY022779-01 to Dr Anne Slavotinek from the National Eye Institute, National Institutes of Health. We thank the California Department of Public Health Maternal Child and Adolescent Health Division for providing data for some analyses. The findings and conclusions in this report are those of the authors and do not necessarily represent the view of the California Department of Public Health.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on European Journal of Human Genetics website
Supplementary information
Rights and permissions
About this article

Cite this article
Choi, A., Lao, R., Ling-Fung Tang, P. et al. Novel mutations in PXDN cause microphthalmia and anterior segment dysgenesis. Eur J Hum Genet 23, 337–341 (2015). https://doi.org/10.1038/ejhg.2014.119
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ejhg.2014.119
This article is cited by
-
Three Novel Mutations of Microphthalmos Identified in Two Chinese Families
Phenomics (2022)
-
CUGC for syndromic microphthalmia including next-generation sequencing-based approaches
European Journal of Human Genetics (2020)
-
Novel PXDN biallelic variants in patients with microphthalmia and anterior segment dysgenesis
Journal of Human Genetics (2020)
-
Unraveling the genetic cause of hereditary ophthalmic disorders in Arab societies from Israel and the Palestinian Authority
European Journal of Human Genetics (2020)
-
Genetics of anophthalmia and microphthalmia. Part 1: Non-syndromic anophthalmia/microphthalmia
Human Genetics (2019)
