
Abstract
Background/Objectives:
Polycystic ovary syndrome (PCOS) is commonly associated with insulin resistance, dyslipidemia and increased inflammation, which all benefit from dietary intake of monounsaturated and n-3 polyunsaturated fatty acids (MUFA and n-3 PUFA). Our goal was to compare the effects of MUFA-rich almonds vs n-3/n-6 PUFA-rich walnuts on metabolic and endocrine parameters in PCOS.
Subjects/Methods:
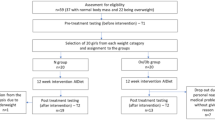
Thirty-one PCOS patients randomly received either walnuts or almonds containing 31 g of total fat per day for 6 weeks. At the beginning and at the end, anthropometric parameters, fasting lipids, phospholipid-fatty acids, inflammatory markers, androgens, oral glucose tolerance tests (OGTT) and frequently sampled intravenous-GTT were obtained.
Results:
Weight remained stable. Within group, walnuts increased the n-3/n-6 essential PUFA in the diet and plasma phospholipids. Walnuts decreased low-density lipoprotein-cholesterol by 6% from 3.76±0.27 to 3.38±0.22 mmol/l (P=0.05) and apoprotein B by 11% from 0.72±0.04 to 0.64±0.05 g/l (P<0.03). Although almonds also reduced low-density lipoprotein-cholesterol by 10% and apoprotein B by 9%, these were not significant. Walnuts increased insulin response during OGTT by 26% (P<0.02). Both walnuts and almonds increased adiponectin (walnuts from 9.5±1.6 to 11.3±1.8 μg per 100 ml, P=0.0241; almonds from 10.1±1.5 to 12.2±1.4 μg/dl, P=0.0262). Walnuts decreased HgBA1 from 5.7±0.1 to 5.5±0.1% (P=0.0006) with significant intergroup difference from almonds (P=0.0470). Walnuts increased sex hormone-binding globulin from 38.3±4.1 to 43.1±4.3 nmol/l (P=0.0038) and almonds reduced free androgen index from 2.6±0.4 to 1.8±0.3 (P=0.0470).
Conclusion:
Nut intake exerted beneficial effects on plasma lipids and androgens in PCOS.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others

References
Abbate R, Gori AM, Martini F, Brunelli T, Filippini M, Francalanci I et al. (1996). n−3 PUFA supplementation, monocyte PCA expression and interleukin-6 production. Prostaglandins Leukot Essent Fatty Acids 54, 439–444.
Almario RU, Vonghavaravat V, Wong R, Kasim-Karakas SE (2001). Effects of walnut consumption on plasma fatty acids and lipoproteins in combined hyperlipidemia. Am J Clin Nutr 74, 72–79.
Arterburn LM, Hall EB, Oken H (2006). Distribution, interconversion, and dose response of n−3 fatty acids in humans. Am J Clin Nutr 83, 1467S–1476S.
Azziz R (2005). Diagnostic criteria for polycystic ovary syndrome: a reappraisal. Fertil steril 83, 1343–1346.
Banel DK, Hu FB (2009). Effects of walnut consumption on blood lipids and other cardiovascular risk factors: a meta-analysis and systematic review. Am J Clin Nutr 90, 56–63.
Baur LA, O’Connor J, Pan DA, Kriketos AD, Storlien LH (1998). The fatty acid composition of skeletal muscle membrane phospholipid: its relationship with the type of feeding and plasma glucose levels in young children. Metabolism 47, 106–112.
Berglund L, Lefevre M, Ginsberg HN, Kris-Etherton PM, Elmer PJ, Stewart PW et al. (2007). Comparison of monounsaturated fat with carbohydrates as a replacement for saturated fat in subjects with a high metabolic risk profile: studies in the fasting and postprandial states. Am J Clin Nutr 86, 1611–1620.
Bonnet F, Balkau B, Malecot JM, Picard P, Lange C, Fumeron F et al. (2009). Sex hormone-binding globulin predicts the incidence of hyperglycemia in women: interactions with adiponectin levels. Eur J Endocrinol 161, 81–85.
Calder PC (2001). Polyunsaturated fatty acids, inflammation, and immunity. Lipids 36, 1007–1024.
Dunaif A (2000). Citation for the 2000 Monsanto Clinical Investigator Award Lecture of The Endocrine Society to Dr William F Crowley. Endocr Rev 21, 449–450.
Dunaif A, Xia J, Book CB, Schenker E, Tang Z (1995). Excessive insulin receptor serine phosphorylation in cultured fibroblasts and in skeletal muscle. A potential mechanism for insulin resistance in the polycystic ovary syndrome. J Clin Invest 96, 801–810.
Ehrmann DA (2005). Polycystic ovary syndrome. N Engl J Med 352, 1223–1236.
Ehrmann DA, Liljenquist DR, Kasza K, Azziz R, Legro RS, Ghazzi MN (2006). Prevalence and predictors of the metabolic syndrome in women with polycystic ovary syndrome. J Clin Endocrinol Metab 91, 48–53.
Feldman EB (2002). The scientific evidence for a beneficial health relationship between walnuts and coronary heart disease. J Nutr 132, 1062S–1101S.
Finnegan YE, Minihane AM, Leigh-Firbank EC, Kew S, Meijer GW, Muggli R et al. (2003). Plant- and marine-derived n−3 polyunsaturated fatty acids have differential effects on fasting and postprandial blood lipid concentrations and on the susceptibility of LDL to oxidative modification in moderately hyperlipidemic subjects. Am J Clin Nutr 77, 783–795.
Glueck CJ, Papanna R, Wang P, Goldenberg N, Sieve-Smith L (2003). Incidence and treatment of metabolic syndrome in newly referred women with confirmed polycystic ovarian syndrome. Metabolism 52, 908–915.
Griel AE, Kris-Etherton PM (2006). Tree nuts and the lipid profile: a review of clinical studies. Br J Nutr 96 (Suppl 2), S68–S78.
Hamilton-Fairley D, White D, Griffiths M, Anyaoku V, Koistinen R, Seppala M et al. (1995). Diurnal variation of sex hormone binding globulin and insulin-like growth factor binding protein-1 in women with polycystic ovary syndrome. Clin Endocrinol (Oxf) 43, 159–165.
Havel PJ (2002). Control of energy homeostasis and insulin action by adipocyte hormones: leptin, acylation stimulating protein, and adiponectin. Curr Opin Lipidol 13, 51–59.
Karakas SE, Kim K, Duleba AJ (2010). Determinants of impaired fasting glucose versus glucose intolerance in polycystic ovary syndrome. Diabetes Care 33, 887–893.
Kasim-Karakas SE, Almario RU, Gregory L, Wong R, Todd H, Lasley BL (2004). Metabolic and endocrine effects of a polyunsaturated fatty acid-rich diet in polycystic ovary syndrome. J Clin Endocrinol Metab 89, 615–620.
Kopecky J, Rossmeisl M, Flachs P, Kuda O, Brauner P, Jilkova Z et al. (2009). n−3 PUFA: bioavailability and modulation of adipose tissue function. Proc Nutr Soc 68, 361–369.
Korotkova M, Gabrielsson B, Lonn M, Hanson LA, Strandvik B (2002). Leptin levels in rat offspring are modified by the ratio of linoleic to alpha-linolenic acid in the maternal diet. J Lipid Res 43, 1743–1749.
Ma QL, Yang F, Rosario ER, Ubeda OJ, Beech W, Gant DJ et al. (2009). Beta-amyloid oligomers induce phosphorylation of tau and inactivation of insulin receptor substrate via c-Jun N-terminal kinase signaling: suppression by omega-3 fatty acids and curcumin. J Neurosci 29, 9078–9089.
Ma Y, Njike VY, Millet J, Dutta S, Doughty K, Treu JA et al. (2010). Effects of walnut consumption on endothelial function in type 2 diabetic subjects: a randomized controlled crossover trial. Diabetes Care 33, 227–232.
Marshall LA, Johnston PV (1982). Modulation of tissue prostaglandin synthesizing capacity by increased ratios of dietary alpha-linolenic acid to linoleic acid. Lipids 17, 905–913.
Martin de Santa Olalla L, Sanchez Muniz FJ, Vaquero MP (2009). N−3 fatty acids in glucose metabolism and insulin sensitivity. Nutr Hosp 24, 113–127.
Pareja A, Tinahones FJ, Soriguer FJ, Monzon A, Esteva de Antonio I, Garcia-Arnes J et al. (1997). Unsaturated fatty acids alter the insulin secretion response of the islets of Langerhans in vitro. Diabetes Res Clin Pract 38, 143–149.
Perez de Heredia F, Sanchez J, Priego T, Larque E, del Puy Portillo M, Palou A et al. (2009). Adiponectin is associated with serum and adipose tissue fatty acid composition in rats. J Endocrinol Invest 32, 659–665.
Peyron-Caso E, Taverna M, Guerre-Millo M, Veronese A, Pacher N, Slama G et al. (2002). Dietary (n−3) polyunsaturated fatty acids up-regulate plasma leptin in insulin-resistant rats. J Nutr 132, 2235–2240.
Reseland JE, Haugen F, Hollung K, Solvoll K, Halvorsen B, Brude IR et al. (2001). Reduction of leptin gene expression by dietary polyunsaturated fatty acids. J Lipid Res 42, 743–750.
Sacks FM, Campos H (2006). Polyunsaturated fatty acids, inflammation, and cardiovascular disease: time to widen our view of the mechanisms. J Clin Endocrinol Metab 91, 398–400.
Schaefer EJ, Lichtenstein AH, Lamon-Fava S, McNamara JR, Schaefer MM, Rasmussen H et al. (1995). Body weight and low-density lipoprotein cholesterol changes after consumption of a low-fat ad libitum diet. JAMA 274, 1450–1455.
Sekine S, Sasanuki S, Murano Y, Aoyama T, Takeuchi H (2008). Alpha-linolenic acid-rich flaxseed oil ingestion increases plasma adiponectin level in rats. Int J Vitam Nutr Res 78, 223–229.
Simopoulos AP, Leaf A, Salem Jr N (2000). Workshop statement on the essentiality of and recommended dietary intakes for Omega-6 and Omega-3 fatty acids. Prostaglandins Leukot Essent Fatty Acids 63, 119–121.
Spranger J, Mohlig M, Wegewitz U, Ristow M, Pfeiffer AF, Schill T et al. (2004). Adiponectin is independently associated with insulin sensitivity in women with polycystic ovary syndrome. Clin Endocrinol (Oxf) 61, 738–746.
Stefan N, Stumvoll M (2002). Adiponectin--its role in metabolism and beyond. Horm Metab Res 34, 469–474.
Takahashi Y, Ide T (2000). Dietary n−3 fatty acids affect mRNA level of brown adipose tissue uncoupling protein 1, and white adipose tissue leptin and glucose transporter 4 in the rat. Br J Nutr 84, 175–184.
Tapsell LC, Batterham MJ, Teuss G, Tan SY, Dalton S, Quick CJ et al. (2009). Long-term effects of increased dietary polyunsaturated fat from walnuts on metabolic parameters in type II diabetes. Eur J Clin Nutr 63, 1008–1015.
Vessby B (2000). Dietary fat and insulin action in humans. Br J nutr 83 (Suppl 1), S91–S96.
Acknowledgements
This study was supported by grants from the National Center for Complementary and Alternative Medicine (R21 AT002280) and the ALSAM Foundation to Dr SE Karakas. The clinical studies were partially supported by the UC Davis Clinical and Translational Science Center grant (RR 024146).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
About this article
Cite this article
Kalgaonkar, S., Almario, R., Gurusinghe, D. et al. Differential effects of walnuts vs almonds on improving metabolic and endocrine parameters in PCOS. Eur J Clin Nutr 65, 386–393 (2011). https://doi.org/10.1038/ejcn.2010.266
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ejcn.2010.266
Keywords
This article is cited by
-
Lifestyle management in polycystic ovary syndrome – beyond diet and physical activity
BMC Endocrine Disorders (2023)
-
Nutritional and health-beneficial values of almond nuts consumption
Nutrire (2023)
-
A comprehensive review of clinical studies with herbal medicine on polycystic ovary syndrome (PCOS)
DARU Journal of Pharmaceutical Sciences (2019)
