
Abstract
Background:
C-reactive protein (CRP), a sensitive marker of inflammation, is an independent predictor of future cardiovascular disease (CVD), which is a major cause of death worldwide. In epidemiological trials, high-fibre intakes have consistently been associated with reduction in CVD risk and CRP levels.
Objective:
The objective of this study was to assess the influence of dietary fibre (DF) on CRP in clinical trials.
Data sources:
Databases were searched from the earliest record to April 2008 and supplemented by crosschecking reference lists of relevant publications.
Study selection:
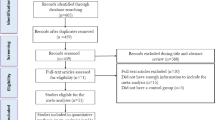
Human adult intervention trials, at least 2 weeks in duration, with an increased and measurable consumption of DF were included and rated for quality.
Data synthesis:
Seven clinical trials were included, and six of these reported significantly lower CRP concentrations of 25–54% with increased DF consumption with dosages ranging between 3.3–7.8 g/MJ. The seventh trial with psyllium fibre supplementation failed to lower CRP levels significantly in overweight/obese individuals. Weight loss and altered fatty acid intakes were present in most of the studies.
Conclusions:
In the presence of weight loss and modified saturated, monounsaturated and polyunsaturated fat intakes, significantly lower CRP concentrations (↓25–54%) are seen with increased fibre consumption (⩾3.3 g/MJ). Mechanisms are inconclusive but may involve the effect of DF on weight loss, and/or changes in the secretion, turnover or metabolism of insulin, glucose, adiponectin, interleukin-6, free fatty acids and triglycerides. Clinical studies of high- and low-fibre diets are needed to explore the potential favourable effects as observed epidemiologically, and to understand individual susceptibility to its anti-inflammatory effect and long-term cardiovascular reduction.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout

Similar content being viewed by others


References
Ajani UA, Ford ES, Mokdad AH (2004). Dietary fiber and C-reactive protein: findings from National Health and Nutrition Examination Survey data. J Nutr 134, 1181–1185.
Anderson JW, Gustafson NJ, Bryant CA, Tietyen-Clark J (1987). Dietary fiber and diabetes: a comprehensive review and practical application. J Am Diet Assoc 87, 1189–1197.
Anderson JW, O’Neal DS, Riddell-Mason S, Floore TL, Dillon DW, Oeltgen PR (1995). Postprandial serum glucose, insulin, and lipoprotein responses to high- and low-fiber diets. Metabolism 44, 848–854.
Anderson JW, Randles KM, Kendall CW, Jenkins DJ (2004). Carbohydrate and fiber recommendations for individuals with diabetes: a quantitative assessment and meta-analysis of the evidence. J Am Coll Nutr 23, 5–17.
Andoh A, Bamba T, Sasaki M (1999). Physiological and anti-inflammatory roles of dietary fiber and butyrate in intestinal functions. J Parenter Enteral Nutr 23, S70–S73.
Arya S, Isharwal S, Misra A, Pandey RM, Rastogi K, Vikram NK et al. (2006). C-reactive protein and dietary nutrients in urban Asian Indian adolescents and young adults. Nutrition 22, 865–871.
Bastard JP, Jardel C, Bruckert E, Blondy P, Capeau J, Laville M et al. (2000). Elevated levels of interleukin 6 are reduced in serum and subcutaneous adipose tissue of obese women after weight loss. J Clin Endocrinol Metab 85, 3338–3342.
Bastard JP, Maachi M, Lagathu C, Kim MJ, Caron M, Vidal H et al. (2006). Recent advances in the relationship between obesity, inflammation, and insulin resistance. Eur Cytokine Netw 17, 4–12.
Basu A, Devaraj S, Jialal I (2006). Dietary factors that promote or retard inflammation. Arterioscler Thromb Vasc Biol 26, 995–1001.
Bemelmans WJ, Lefrandt JD, Feskens EJ, van Haelst PL, Broer J, Meyboom-de Jong B et al. (2004). Increased alpha-linolenic acid intake lowers C-reactive protein, but has no effect on markers of atherosclerosis. Eur J Clin Nutr 58, 1083–1089.
Berg AH, Scherer PE (2005). Adipose tissue, inflammation, and cardiovascular disease. Circ Res 96, 939–949.
Blankenberg S, Tiret L, Bickel C, Peetz D, Cambien F, Meyer J et al. (2002). Interleukin-18 is a strong predictor of cardiovascular death in stable and unstable angina. Circulation 106, 24–30.
Bo S, Durazzo M, Guidi S, Carello M, Sacerdote C, Silli B et al. (2006). Dietary magnesium and fiber intakes and inflammatory and metabolic indicators in middle-aged subjects from a population-based cohort. Am J Clin Nutr 84, 1062–1069.
Borges RL, Ribeiro-Filho FF, Carvalho MB, Zanella MT (2007). Impact of weight loss on adipocytokines, C-reactive protein and insulin sensitivity in hypertensive women with central obesity. Arq Bras Cardiol 89, 371–375.
Burton-Freeman B, Davis PA, Schneeman BO (2002). Plasma cholecystokinin is associated with subjective measures of satiety in women. Am J Clin Nutr 76, 659–667.
Castell JV, Gomez-Lechon MJ, David M, Andus T, Geiger T, Trullenque R et al. (1989). Interleukin-6 is the major regulator of acute phase protein synthesis in adult human hepatocytes. FEBS Lett 242, 237–239.
Chambers JC, Eda S, Bassett P, Karim Y, Thompson SG, Gallimore JR et al. (2001). C-reactive protein, insulin resistance, central obesity, and coronary heart disease risk in Indian Asians from the United Kingdom compared with European whites. Circulation 104, 145–150.
Chandalia M, Garg A, Lutjohann D, von Bergmann K, Grundy SM, Brinkley LJ (2000). Beneficial effects of high dietary fiber intake in patients with type 2 diabetes mellitus. N Engl J Med 342, 1392–1398.
Clifton PM (2003). Diet and C-reactive protein. Curr Atheroscler Rep 5, 431–436.
Cummings JH, Englyst HN (1987). Fermentation in the human large intestine and the available substrates. Am J Clin Nutr 45, 1243–1255.
Dvorakova-Lorenzova A, Suchanek P, Havel PJ, Stavek P, Karasova L, Valenta Z et al. (2006). The decrease in C-reactive protein concentration after diet and physical activity induced weight reduction is associated with changes in plasma lipids, but not interleukin-6 or adiponectin. Metabolism 55, 359–365.
Esposito K, Marfella R, Ciotola M, Di Palo C, Giugliano F, Giugliano G et al. (2004). Effect of a Mediterranean-style diet on endothelial dysfunction and markers of vascular inflammation in the metabolic syndrome—a randomized trial. J Am Med Assoc 292, 1440–1446.
Esposito K, Nappo F, Giugliano F, Di Palo C, Ciotola M, Barbieri M et al. (2003b). Meal modulation of circulating interleukin 18 and adiponectin concentrations in healthy subjects and in patients with type 2 diabetes mellitus. Am J Clin Nutr 78, 1135–1140.
Esposito K, Nappo F, Marfella R, Giugliano G, Giugliano F, Ciotola M et al. (2002). Inflammatory cytokine concentrations are acutely increased by hyperglycemia in humans: role of oxidative stress. Circulation 106, 2067–2072.
Esposito K, Pontillo A, Di Palo C, Giugliano G, Masella M, Marfella R et al. (2003a). Effect of weight loss and lifestyle changes on vascular inflammatory markers in obese women: a randomized trial. J Am Med Assoc 289, 1799–1804.
Esposito K, Giugliano D (2006). Whole-grain intake cools down inflammation. Am J Clin Nutr 83: 1440–1441.
Fain JN, Madan AK, Hiler ML, Cheema P, Bahouth SW (2004). Comparison of the release of adipokines by adipose tissue, adipose tissue matrix, and adipocytes from visceral and subcutaneous abdominal adipose tissues of obese humans. Endocrinology 145, 2273–2282.
Fasshauer M, Klein J, Lossner U, Paschke R (2003). Interleukin (IL)-6 mRNA expression is stimulated by insulin, isoproterenol, tumour necrosis factor alpha, growth hormone, and IL-6 in 3T3-L1 adipocytes. Horm Metab Res 35, 147–152.
Ferchaud-Roucher V, Pouteau E, Piloquet H, Zair Y, Krempf M (2005). Colonic fermentation from lactulose inhibits lipolysis in overweight subjects. Am J Physiol Endocrinol Metab 289, E716–E720.
Fernandez-Real JM, Ricart W (2003). Insulin resistance and chronic cardiovascular inflammatory syndrome. Endocr Rev 24, 278–301.
Flight I, Clifton P (2006). Cereal grains and legumes in the prevention of coronary heart disease and stroke: a review of the literature. Eur J Clin Nutr 60, 1145–1159.
Florez H, Castillo-Florez S, Mendez A, Casanova-Romero P, Larreal-Urdaneta C, Lee D et al. (2006). C-reactive protein is elevated in obese patients with the metabolic syndrome. Diabetes Res Clin Pract 71, 92–100.
Fogarty AW, Glancy C, Jones S, Lewis SA, McKeever TM, Britton JR (2008). A prospective study of weight change and systemic inflammation over 9 y. Am J Clin Nutr 87, 30–35.
Ford ES (2003). The metabolic syndrome and C-reactive protein, fibrinogen, and leukocyte count: findings from the Third National Health and Nutrition Examination Survey. Atherosclerosis 168, 351–358.
Fredrikson GN, Hedblad B, Nilsson JA, Alm R, Berglund G, Nilsson J (2004). Association between diet, lifestyle, metabolic cardiovascular risk factors, and plasma C-reactive protein levels. Metabolism 53, 1436–1442.
Havel PJ (2004). Update on adipocyte hormones: regulation of energy balance and carbohydrate/lipid metabolism. Diabetes 53 (Suppl 1), S143–S151.
Holt S, Brand J, Soveny C, Hansky J (1992). Relationship of satiety to postprandial glycaemic, insulin and cholecystokinin responses. Appetite 18, 129–141.
Howarth NC, Saltzman E, Roberts SB (2001). Dietary fiber and weight regulation. Nutr Rev 59, 129–139.
Iwaki M, Matsuda M, Maeda N, Funahashi T, Matsuzawa Y, Makishima M et al. (2003). Induction of adiponectin, a fat-derived antidiabetic and antiatherogenic factor, by nuclear receptors. Diabetes 52, 1655–1663.
Jenkins DJ, Kendall CW, Marchie A, Faulkner DA, Wong JM, de Souza R et al. (2003). Effects of a dietary portfolio of cholesterol-lowering foods vs lovastatin on serum lipids and C-reactive protein. J Am Med Assoc 290, 502–510.
Jenkins DJ, Kendall CW, Popovich DG, Vidgen E, Mehling CC, Vuksan V et al. (2001). Effect of a very-high-fiber vegetable, fruit, and nut diet on serum lipids and colonic function. Metabolism 50, 494–503.
Junker R, Pieke B, Schulte H, Nofer R, Neufeld M, Assmann G et al. (2001). Changes in hemostasis during treatment of hypertriglyceridemia with a diet rich in monounsaturated and n-3 polyunsaturated fatty acids in comparison with a low-fat diet. Thromb Res 101, 355–366.
Kanauchi O, Mitsuyama K, Araki Y, Andoh A (2003). Modification of intestinal flora in the treatment of inflammatory bowel disease. Curr Pharm Des 9, 333–346.
Kerner A, Avizohar O, Sella R, Bartha P, Zinder O, Markiewicz W et al. (2005). Association between elevated liver enzymes and C-reactive protein: possible hepatic contribution to systemic inflammation in the metabolic syndrome. Arterioscler Thromb Vasc Biol 25, 193–197.
Khera A, McGuire DK, Murphy SA, Stanek HG, Das SR, Vongpatanasin W et al. (2005). Race and gender diffences in C-reactive protein levels. J Am Coll Cardiol 46, 464–469.
King DE (2005). Dietary fiber, inflammation, and cardiovascular disease. Mol Nutr Food Res 49, 594–600.
King DE, Egan BM, Geesey ME (2003). Relation of dietary fat and fiber to elevation of C-reactive protein. Am J Cardiol 92, 1335–1339.
King DE, Egan BM, Woolson RF, Mainous AG, Al-Solaiman Y, Jesri A (2007). Effect of a high-fiber diet vs a fiber-supplemented diet on C-reactive protein level. Arch Intern Med 167, 502–506.
King DE, Mainous AG, Egan BM, Woolson RF, Geesey ME (2008). Effect of psyllium fiber supplementation on C-reactive protein: the trial to reduce inflammatory markers (TRIM). Ann Fam Med 6, 100–106.
Klein S, Fontana L, Young VL, Coggan AR, Kilo C, Patterson BW et al. (2004). Absence of an effect of liposuction on insulin action and risk factors for coronary heart disease. N Engl J Med 350, 2549–2557.
Koren MS, Purnell JQ, Breen PA, Matthys CC, Callahan HS, Weigle DS (2006). Plasma C-reactive protein concentration is not affected by isocaloric dietary fat reduction. Nutrition 22, 444–448.
Krogh-Madsen R, Plomgaard P, Keller P, Keller C, Pedersen BK (2004). Insulin stimulates interleukin-6 and tumor necrosis factor-alpha gene expression in human subcutaneous adipose tissue. Am J Physiol 286, E234–E238.
Liu S, Manson JE, Buring JE, Stampfer MJ, Willett WC, Ridker PM (2002). Relation between a diet with a high glycemic load and plasma concentrations of high-sensitivity C-reactive protein in middle-aged women. Am J Clin Nutr 75, 492–498.
Lopez-Garcia E, Schulze MB, Manson JE, Meigs JB, Albert CM, Rifai N et al. (2004). Consumption of (n-3) fatty acids is related to plasma biomarkers of inflammation and endothelial activation in women. J Nutr 134, 1806–1811.
Ma Y, Griffith JA, Chasan-Taber L, Olendzki BC, Jackson E, Stanek III EJ et al. (2006). Association between dietary fiber and serum C-reactive protein. Am J Clin Nutr 83, 760–766.
Ma Y, Hebert JR, Li W, Bertone-Johnson ER, Olendzki BC, Pagoto SL et al. (2008). Association between dietary fiber and markers of systemic inflammation in the Women's Health Initiative Observational Study. Nutrition 24, 941–942.
MacGregor AJ, Gallimore JR, Spector TD, Pepys MB (2004). Genetic effects on baseline values of C-reactive protein and serum amyloid a protein: a comparison of monozygotic and dizygotic twins. Clin Chem 50, 130–134.
Marckmann P, Sandström B, Jespersen J (1994). Low-fat, high-fiber diet favorably affects several independent risk markers of ischemic heart disease: observations on blood lipids, coagulation, and fibrinolysis from a trial of middle-aged Danes. Am J Clin Nutr 59, 935–939.
Marfella R, Esposito K, Siniscalchi M, Cacciapuoti F, Giugliano F, Labriola D et al. (2004). Effect of weight loss on cardiac synchronization and proinflammatory cytokines in premenopausal obese women. Diabetes Care 27, 47–52.
McLaughlin T, Abbasi F, Lamendola C, Liang L, Reaven G, Schaaf P et al. (2002). Differentiation between obesity and insulin resistance in the association with C-reactive protein. Circulation 106, 2908–2912.
Mohamed-Ali V, Goodrick S, Rawesh A, Katz DR, Miles JM, Yudkin JS et al. (1997). Subcutaneous adipose tissue releases interleukin-6, but not tumor necrosis factor-alpha, in vivo. J Clin Endocrinol Metab 82, 4196–4200.
O'Brien KD, Brehm BJ, Seeley RJ, Bean J, Wener MH, Daniels S et al. (2005). Diet-induced weight loss is associated with decreases in plasma serum amyloid a and C-reactive protein independent of dietary macronutrient composition in obese subjects. J Clin Endocrinol Metab 90, 2244–2249.
Parks EJ, Krauss RM, Christiansen MP, Neese RA, Hellerstein MK (1999). Effects of a low-fat, high-carbohydrate diet on VLDL-triglyceride assembly, production, and clearance. J Clin Invest 104, 1087–1096.
Pearson TA, Mensah GA, Alexander RW, Anderson JL, Cannon III RO, Criqui M et al. (2003). Markers of inflammation and cardiovascular disease: application to clinical and public health practice: a statement for healthcare professionals from the Centers for Disease Control and Prevention and the American Heart Association. Circulation 107, 499–511.
Pepys MB, Hirschfield GM (2003). C-reactive protein: a critical update. J Clin Invest 111, 1805–1812.
Pereira MA, Ludwig DS (2001). Dietary fiber and body-weight regulation. Observations and mechanisms. Pediatr Clin North Am 48, 969–980.
Pereira MA, O’Reilly E, Augustsson K, Fraser GE, Goldbourt U, Heitman BL et al. (2004). Dietary fiber and risk of coronary heart disease: a pooled analysis of cohort studies. Arch Intern Med 164, 370–376.
Qi L, Rimm E, Liu S, Rifai N, Hu FB (2005). Dietary glycemic index, glycemic load, cereal fiber, and plasma adiponectin concentration in diabetic men. Diabetes Care 28, 1022–1028.
Qi L, van Dam RM, Liu S, Franz M, Mantzoros C, Hu FB (2006). Whole-grain, bran, and cereal fiber intakes and markers of systemic inflammation in diabetic women. Diabetes Care 29, 207–211.
Rallidis LS, Paschos G, Liakos GK, Velissaridou AH, Anastasiadis G, Zampelas A (2003). Dietary alpha-linolenic acid decreases C-reactive protein, serum amyloid A and interleukin-6 in dyslipidaemic patients. Atherosclerosis 167, 237–242.
Rezar V, Pajk T, Marinsek LR, Jese JV, Salobir K, Oresnik A et al. (2003). Wheat bran and oat bran effectively reduce oxidative stress induced by high-fat diets in pigs. Ann Nutr Metab 47, 78–84.
Richter V, Purschwitz K, Rassoul F, Thiery J, Zunft HJ, Leitzmann C (2004). Effects of diet modification on cardiovascular risk: results from the Leipzig Wholesome Nutrition study. Asia Pac J Clin Nutr 13, S106.
Ridker PM (2005). C-reactive protein, inflammation, and cardiovascular disease: clinical update. Tex Heart Inst J 32, 384–386.
Ridker PM, Buring JE, Shih J, Matias M, Henneken SC (1998). Prospective study of C-reactive protein and the risk of future cardiovascular events among apparently healthy women. Circulation 98, 731–733.
Ridker PM, Rifai N, Rose L, Buring JE, Cook NR (2002). Comparison of C-reactive and low density lipoprotein cholesterol levels in the prediction of first cardiovascular events. N Engl J Med 347, 1557–1565.
Rivellese AA, Auletta P, Marotta G, Saldalamacchia G, Giacco A, Mastrilli V et al. (1994). Long term metabolic effects of two dietary methods of treating hyperlipidaemia. BMJ 308, 227–231.
Rosamond WD (2002). Dietary fiber and prevention of cardiovascular disease. J Am Coll Cardiol 39, 57–59.
Salmeron J, Manson JE, Stampfer MJ, Colditz GA, Wing AL, Willett WC (1997). Dietary fiber, glycemic load, and risk of non-insulin-dependent diabetes mellitus in women. J Am Med Assoc 277, 472–477.
Saris WH, Astrup A, Prentice AM, Zunft HJ, Formiguera X, Verboeket-van de Venne WP et al. (2000). Randomized controlled trial of changes in dietary carbohydrate/fat ratio and simple vs complex carbohydrates on body weight and blood lipids: the CARMEN study. The Carbohydrate Ratio Management in European National diets. Int J Obes Relat Metab Disord 24, 1310–1318.
Scheppach W, Wiggins HS, Halliday D, Self R, Howard J, Branch WJ et al. (1988). Effect of gut-derived acetate on glucose turnover in man. Clin Sci 75, 363–370.
Schulze MB, Liu S, Rimm EB, Manson JE, Willett WC, Hu FB (2004). Glycemic index, glycemic load, and dietary fiber intake and incidence of type 2 diabetes in younger and middle-aged women. Am J Clin Nutr 80, 348–356.
Selvin E, Paynter NP, Erlinger TP (2007). The effect of weight loss on C-reactive protein. A systematic review. Arch Intern Med 167, 31–39.
Shigematsu R, Okura T, Kumagai S, Kai Y, Hiyama T, Sasaki H et al. (2006). Cutoff and target values for intra-abdominal fat area for prevention of metabolic disorders in pre- and post-menopausal obese women before and after weight reduction. Circ J 70, 110–114.
Slavin JL (2005). Dietary fiber and body weight. Nutrition 21, 411–418.
Taskinen MR (1990). Hyperlipidaemia in diabetes. Baillieres Clin Endocrinol Metab 4, 743–775.
Trowell HC (1975). Dietary-fiber hypothesis of the etiology of diabetes mellitus. Diabetes 24, 762–765.
Tschritter O, Fritsche A, Thamer C, Haap M, Shirkavand F, Rahe S et al. (2003). Plasma adiponectin concentrations predict insulin sensitivity of both glucose and lipid metabolism. Diabetes 52, 239–243.
Verhagen AP, de Vet HC, de Bie RA, Kessels AG, Boers M, Bouter LM et al. (1998). The Delphi list: a criteria list for quality assessment of randomized clinical trials for conducting systematic reviews developed by Delphi consensus. J Clin Epidemiol 51, 1235–1241.
Vicennati V, Vottero A, Friedman C, Papanicolaou DA (2002). Hormonal regulation of interleukin-6 production in human adipocytes. Int J Obes Relat Metab Disord 26, 905–911.
Vigushin DM, Pepys MB, Hawkins PN (1993). Metabolic and scintigraphic studies of radioiodinated human C-reactive protein in health and disease. J Clin Invest 91, 1351–1357.
Weickert MO, Mohlig M, Schofl C, Arafat AM, Otto B, Viehoff H et al. (2006). Cereal fiber improves whole-body insulin sensitivity in overweight and obese women. Diabetes Care 29, 775–780.
Yamauchi T, Kamon J, Waki H, Terauchi Y, Kubota N, Hara K et al. (2001). The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat Med 7, 941–946.
Yao M, Roberts SB (2001). Dietary energy density and weight regulation. Nutr Rev 59, 247–258.
Yudkin JS, Stehouwer CD, Emeis JJ, Coppack SW (1999). C-reactive protein in healthy subjects: associations with obesity, insulin resistance, and endothelial dysfunction: a potential role for cytokines originating from adipose tissue? Arterioscler Thromb Vasc Biol 19, 972–978.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
North, C., Venter, C. & Jerling, J. The effects of dietary fibre on C-reactive protein, an inflammation marker predicting cardiovascular disease. Eur J Clin Nutr 63, 921–933 (2009). https://doi.org/10.1038/ejcn.2009.8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ejcn.2009.8
Keywords
This article is cited by
-
The association between macronutrient intakes and coronavirus disease 2019 (COVID-19) in an Iranian population: applying a dynamical system model
Journal of Health, Population and Nutrition (2023)
-
The effect of DASH diet on glycemic response, meta-inflammation and serum LPS in obese patients with NAFLD: a double-blind controlled randomized clinical trial
Nutrition & Metabolism (2023)
-
Association between dietary fiber intake and atherosclerotic cardiovascular disease risk in adults: a cross-sectional study of 14,947 population based on the National Health and Nutrition Examination Surveys
BMC Public Health (2022)
-
Host-microbiome interactions: Gut-Liver axis and its connection with other organs
npj Biofilms and Microbiomes (2022)
-
Association between plant-based diets and metabolic health status in adolescents with overweight and obesity
Scientific Reports (2022)
