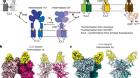
- NEWS AND VIEWS
Hit movie reveals how a tuberculosis drug halts ATP synthesis
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 589, 21-22 (2021)
doi: https://doi.org/10.1038/d41586-020-03406-x
References
Guo, H. et al. Nature 589, 143–147 (2021).
Schnippel, K. et al. Lancet Respir. Med. 6, 699–706 (2018).
Conradie, F. et al. N. Engl. J. Med. 382, 893–902 (2020).
Andries, K. et al. Science 307, 223–227 (2005).
Kühlbrandt, W. Annu. Rev. Biochem. 88, 515–549 (2019).
Roh, S.-H. et al. Sci. Adv. 6, eabb9605 (2020).
Haagsma, A. C., Driessen, N. N., Hahn, M.-M., Lill, H. & Bald, D. FEMS Microbiol. Lett. 313, 68–74 (2010).
Zhang, A. T., Montgomery, M. G., Leslie, A. G. W., Cook, G. M. & Walker, J. E. Proc. Natl Acad. Sci. USA 116, 4206–4211 (2019).
Lu, P., Lill, H. & Bald, D. Biochim. Biophys. Acta 1837, 1208–1218 (2014).
Ragunathan, P. et al. J. Biol. Chem. 292, 11262–11279 (2017).
Koul, A. et al. Nature Chem. Biol. 3, 323–324 (2007).
Preiss, L. et al. Sci. Adv. 1, e1500106 (2015).
 Read the paper: Structure of mycobacterial ATP synthase bound to the tuberculosis drug bedaquiline
Read the paper: Structure of mycobacterial ATP synthase bound to the tuberculosis drug bedaquiline
 Tuberculosis vaccine finds an improved route
Tuberculosis vaccine finds an improved route
 Drug discovery goes au naturel
Drug discovery goes au naturel