- NEWS AND VIEWS
Prioritizing where to restore Earth’s ecosystems
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 586, 680-681 (2020)
doi: https://doi.org/10.1038/d41586-020-02750-2
References
Temperton, V. M. et al. Restor. Ecol. 27, 705–719 (2019).
Strassburg, B. N. et al. Nature 586, 724–729 (2020).
Convention on Biological Diversity. COP Decision X/2: Strategic Plan for Biodiversity 2011–2020 (2012).
Dinerstein, E. et al. Sci. Adv. 6, eabb2824 (2020).
Pecl, G. T. et al. Science 355, eaai9214 (2017).
Di Marco, M. et al. Glob. Change Biol. 25, 2763–2778 (2019).
Beyer, H. L., Venter, O., Grantham, H. S. & Watson, J. E. M. Conserv. Lett. 13, e12692 (2020).
Maron, M. et al. Nature Ecol. Evol. 4, 46–49 (2020).
Leclère, D. et al. Nature 585, 551–556 (2020).
Nicholson, E. et al. Trends Ecol. Evol. 34, 57–68 (2019).
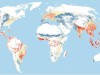 Read the paper: Global priority areas for ecosystem restoration
Read the paper: Global priority areas for ecosystem restoration
 A recipe to reverse the loss of nature
A recipe to reverse the loss of nature
 Rethinking extinctions that arise from habitat loss
Rethinking extinctions that arise from habitat loss