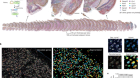
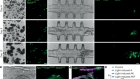
- TECHNOLOGY FEATURE
Technologies to watch in 2020
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 577, 585-587 (2020)
doi: https://doi.org/10.1038/d41586-020-00114-4
These interviews have been edited for length and clarity.
References
Liu, N. et al. J. Am. Chem. Soc. 141, 4016–4025 (2019).
Wei, H. et al. J. Struct. Biol. 202, 170–174 (2018).
Schaffer, M. et al. Nature Meth.16, 757–762 (2019).
Noecker, C. et al. mSystems https://doi.org/10.1128/msystems.00579-19 (2019).
Chan, M. M. et al. Nature 570, 77–82 (2019).
Kalhor, R. et al. Science 361, eaat9804 (2018).
Askary, A. et al. Nature Biotechnol. 38, 66–75 (2020).
Hu, Z. et al. Nature Genet. 51, 1113–1122 (2019).
Mich, J. et al. Preprint at bioRxiv https://doi.org/10.1101/555318 (2019).
Hrvatin, S. et al. eLife 8, e48089 (2019).
Colasante, G. et al. Mol. Ther. https://doi.org/10.1016/j.ymthe.2019.08.018 (2019).
Gierahn, T. M. et al. Nature Meth. 14, 395–398 (2017).
Hughes, T. K. et al. Preprint at bioRxiv https://doi.org/10.1101/689273 (2019).
Tu, A. A. et al. Nature Immunol. 20, 1692–1699 (2019).
Kula, T. et al. Cell 178, 1016–1028 (2019).
Wang, H. et al. Cell 175, 1405–1417 (2018).
Kim, J. H. et al. Nature Meth. 16, 633–639 (2019).
Chin, Y. et al. Cell 175, 1481–1491 (2018).
 NatureTech
NatureTech
 Single-cell approaches to immune profiling
Single-cell approaches to immune profiling
 Microbial chemistry gains fresh focus
Microbial chemistry gains fresh focus
 Technologies to watch in 2019
Technologies to watch in 2019