- NEWS AND VIEWS
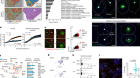
New predictors for immunotherapy responses sharpen our view of the tumour microenvironment
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 577, 474-476 (2020)
doi: https://doi.org/10.1038/d41586-019-03943-0
References
Brahmer, J. R. et al. J. Clin. Oncol. 28, 3167–3175 (2010).
Cabrita, R. et al. Nature 577, 561–565 (2020).
Petitprez, F. et al. Nature 577, 556–560 (2020).
Helmink, B. A. et al. Nature https://doi.org/10.1038/s41586-019-1922-8 (2020).
Shimabukuro-Vornhagen, A. et al. Oncotarget 5, 4651–4664 (2014).
Germain, C. et al. Am. J. Respir. Crit. Care Med. 189, 832–844 (2014).
Shalapour, S. et al. Nature 521, 94–98 (2015).
DeFalco, J. et al. Clin. Immunol. 187, 37–45 (2018).
Bruno, T. C. et al. Cancer Immunol. Res. 5, 898–907 (2017).
Kessel, A. et al. Autoimmun. Rev. 11, 670–677 (2012).
Khan, A. R. et al. Nature Commun. 6, 5997 (2015).
Sautès-Fridman, C., Petitprez, F., Calderaro, J. & Fridman, W. H. Nature Rev. Cancer 19, 307–325 (2019).
Affara, N. I. et al. Cancer Cell 25, 809–821 (2014).
Shalapour, S. et al. Nature 551, 340–345 (2017).
Ammirante, M. et al. Nature 464, 302–305 (2010).
 Read the paper: B cells and tertiary lymphoid structures promote immunotherapy response
Read the paper: B cells and tertiary lymphoid structures promote immunotherapy response
 Read the paper: B cells are associated with survival and immunotherapy response in sarcoma
Read the paper: B cells are associated with survival and immunotherapy response in sarcoma
 Read the paper: Tertiary lymphoid structures improve immunotherapy and survival in melanoma
Read the paper: Tertiary lymphoid structures improve immunotherapy and survival in melanoma
 Teamwork by different T-cell types boosts tumour destruction by immunotherapy
Teamwork by different T-cell types boosts tumour destruction by immunotherapy
 Trispecific antibodies offer a third way forward for anticancer immunotherapy
Trispecific antibodies offer a third way forward for anticancer immunotherapy