- NEWS AND VIEWS
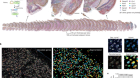
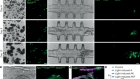
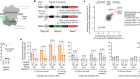
CRISPR tool modifies genes precisely by copying RNA into the genome
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 576, 48-49 (2019)
doi: https://doi.org/10.1038/d41586-019-03392-9
References
Anzalone, A. V. et al. Nature 576, 149–157 (2019).
Matsuoka, Y. et al. Proc. Natl Acad. Sci. USA 99, 6080–6084 (2002).
Capecchi, M. R. Science 244, 1288–1292 (1989).
Rudin, N., Sugarman, E. & Haber, J. E. Genetics 122, 519–534 (1989).
Rouet, P., Smih, F. & Jasin, M. Mol. Cell. Biol. 14, 8096–8106 (1994).
Jinek, M. et al. Science 337, 816–821 (2012).
Cong, L. et al. Science 339, 819–823 (2013).
Mali, P. et al. Science 339, 823–826 (2013).
Komor, A. C., Kim, Y. B., Packer, M. S., Zuris, J. A. & Liu, D. R. Nature 533, 420–424 (2016).
Keskin, H. et al. Nature 515, 436–439 (2014).
Platt, R. J. et al. Cell Rep. 19, 335–350 (2017).
Cox, D. B. T., Platt, R. J. & Zhang, F. Nature Med. 21, 121–131 (2015).
 Read the paper: Search-and-replace genome editing without double-strand breaks or donor DNA
Read the paper: Search-and-replace genome editing without double-strand breaks or donor DNA
 Hijack of CRISPR defences by selfish genes holds clinical promise
Hijack of CRISPR defences by selfish genes holds clinical promise
 On the road to a gene drive in mammals
On the road to a gene drive in mammals