- NEWS AND VIEWS
How plants perceive salt
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 572, 318-320 (2019)
doi: https://doi.org/10.1038/d41586-019-02289-x
References
Schroeder, J. I. et al. Nature 497, 60–66 (2013).
Jiang, Z. et al. Nature 572, 341–346(2019).
Choi, W. G., Toyota, M., Kim, S. H., Hilleary, R. & Gilroy, S. Proc. Natl Acad. Sci. USA 111, 6497–6502 (2014).
Steinhorst, L. & Kudla, J. Curr. Opin. Plant Biol. 22, 14–21 (2014).
Yang, Y. & Guo, Y. New Phytol. 217, 523–539 (2018).
Kim, B. G. et al. Plant J. 52, 473–484 (2007).
Cacas, J. L. et al. Plant Physiol. 170, 367–384 (2016).
Tartaglio, V. et al. Plant J. 89, 278–290 (2017).
Posse de Chaves, E. & Sipione, S. FEBS Lett. 584, 1748–1759 (2010).
Manishankar, P., Wang, N., Köster, P., Alatar, A. A. & Kudla, J. J. Exp. Bot. 69, 4215–4226 (2018).
Pei, Z. M. et al. Nature 406, 731–734 (2000).
Nagano, M. et al. Plant Cell 28, 1966–1983 (2016).
Hao, H. et al. Plant Cell 26, 1729–1745 (2014).
Platre, M. P. et al. Science 364, 57–62 (2019).
Lenarčič, T. et al. Science 358, 1431–1434 (2017).
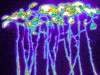 Read the paper: Plant cell-surface GIPC sphingolipids sense salt to trigger Ca2+ influx
Read the paper: Plant cell-surface GIPC sphingolipids sense salt to trigger Ca2+ influx
 Peptide signal alerts plants to drought
Peptide signal alerts plants to drought
 Sexual attraction channelled in moss
Sexual attraction channelled in moss