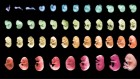- NEWS AND VIEWS
A deep dive into the development of sea squirts
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 571, 333-334 (2019)
doi: https://doi.org/10.1038/d41586-019-01967-0
References
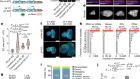
Cao, C. et al. Nature 571, 349–354 (2019).
Stuart, T. & Satija, R. Nature Rev. Genet. 20, 257–272 (2019).
Farrell, J. A. et al. Science 360, eaar3131 (2018).
Briggs, J. A. et al. Science 360, eaar5780 (2018).
Conklin, E. G. J. Acad. Nat. Sci. Philadelphia 13, 1–119 (1905).
Dehal, P. et al. Science 298, 2157–2167 (2002).
Imai, K. S., Levine, M., Satoh, N. & Satou, Y. Science 312, 1183–1187 (2006).
Satoh, N. Developmental Genomics of Ascidians (Wiley-Blackwell, 2014).
Harder, M., Reeves, W., Byers, C., Santiago, M. & Veeman, M. Dev. Biol. 448, 136–146 (2019).
Gans, C. & Northcutt, R. G. Science 220, 268–273 (1983).
Stolfi, A., Ryan, K., Meinertzhagen, I. A. & Christiaen, L. Nature 527, 371–374 (2015).
Horie, R. et al. Nature 560, 228–232 (2018).
Satoh, N. Chordate Origins and Evolution (Academic, 2016).
Gee, H. Across the Bridge: Understanding the Origin of the Vertebrates (Univ. Chicago Press, 2018).
Inoue, J. & Satoh, N. Mol. Biol. Evol. 35, 914–924 (2018).
 Read the paper: Comprehensive single-cell transcriptome lineages of a proto-vertebrate
Read the paper: Comprehensive single-cell transcriptome lineages of a proto-vertebrate
 Acorn worms in a nutshell
Acorn worms in a nutshell
 On the crest of becoming vertebrate
On the crest of becoming vertebrate