- NEWS AND VIEWS
Star-like cells spark behavioural hyperactivity in mice
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 571, 43-44 (2019)
doi: https://doi.org/10.1038/d41586-019-01949-2
References
Clarke, L. E. & Barres, B. A. Nature Rev. Neurosci. 14, 311–321 (2013)
Khakh, B. S. & Sofroniew, M. V. Nature Neurosci. 18, 942–952 (2015).
Araque, A. et al. Neuron 81, 728–739 (2014).
Attwell, D. et al. Nature 468, 232–243 (2010).
Cui, Y. et al. Nature 554, 323–327 (2018).
Adamsky, A. et al. Cell 174, 59–71 (2018).
Gomez, J. A. et al. Nature Commun. 10, 1455 (2019).
Robin, L. M. et al. Neuron 98, 935–944 (2018).
Tong, X. et al. Nature Neurosci. 17, 694–703 (2014).
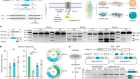
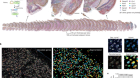
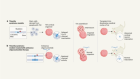
Nagai, J. et al. Cell 177, 1280–1292 (2019).
Klaus, A., da Silva, J. A. & Costa, R. M. Annu. Rev. Neurosci. https://doi.org/10.1146/annurev-neuro-072116-031033 (2019).
Thapar, A. & Cooper, M. Lancet 387, 1240–1250 (2016).
Durston, S., van Belle, J. & de Zeeuw, P. Biol. Psychiatry 69, 1178–1184 (2011).
Kreitzer, A. C. & Malenka, R. C. Neuron 60, 543–554 (2008).
Chai, H. et al. Neuron 95, 531–549 (2017).
Christopherson, K. S. et al. Cell 120, 421–433 (2005).
Bazargani, N. & Attwell, D. Nature Neurosci. 19, 182–189 (2016).
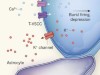 Burst firing sets the stage for depression
Burst firing sets the stage for depression
 Diversity reaches the stars
Diversity reaches the stars