- NEWS AND VIEWS
How an activation signal is transmitted through an excitatory receptor
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 566, 42-43 (2019)
doi: https://doi.org/10.1038/d41586-018-07885-x
References
Koehl, A. et al. Nature 566, 79–84 (2019).
Hauser, A. S., Attwood, M. M., Rask-Andersen, M., Schiöth, H. B. & Gloriam, D. E. Nature Rev. Drug Discov. 16, 829–842 (2017).
Dobrovetsky, E. et al. Protein Data Bank: 3LMK. https://www.rcsb.org/structure/3LMK (2010).
El Moustaine, D. et al. Proc. Natl Acad. Sci. USA 109, 16342–16347 (2012).
Xue, L. et al. Nature Chem. Biol. 11, 134–140 (2015).
Huang, S. et al. Proc. Natl Acad. Sci. USA 108, 15480–15485 (2011).
Liu, J. et al. eLife 6, e26985 (2017).
Wu, H. et al. Science 344, 58–64 (2014).
Wheatley, M. et al. Br. J. Pharmacol. 165, 1688–1703 (2011).
Doré, A. S. et al. Nature 511, 557–562 (2014).
Christopher, J. A. et al. J. Med. Chem. 58, 6653–6664 (2015).
Christopher, J. A. et al. J. Med. Chem. https://doi.org/10.1021/acs.jmedchem.7b01722 (2019).
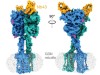 Read the paper: Structural insights into the activation of metabotropic glutamate receptors
Read the paper: Structural insights into the activation of metabotropic glutamate receptors
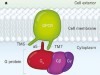 How the ubiquitous GPCR receptor family selectively activates signalling pathways
How the ubiquitous GPCR receptor family selectively activates signalling pathways
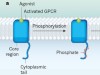 Activation mechanisms for a universal signalling protein
Activation mechanisms for a universal signalling protein