- NEWS AND VIEWS
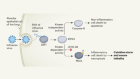
Bacterial defence molecules target viral DNA
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 564, 199-200 (2018)
doi: https://doi.org/10.1038/d41586-018-07576-7
References
Kronheim, S. et al. Nature 564, 283–286 (2018).
Clokie, M. R., Millard, A. D., Letarov, A. V. & Heaphy, S. Bacteriophage 1, 31–45 (2011).
Makarova, K. S. et al. Nature Rev. Microbiol. 12, 722–736 (2015).
Labrie, S. J., Samson, J. E. & Moineau, S. Nature Rev. Microbiol. 8, 317–327 (2010).
Doron, S. et al. Science 359, eaar4120 (2108).
Manning, A. J. & Kuehn, M. J. BMC Microbiol. 1, 258 (2011).
Bamford, D. H. Res. Microbiol. 154, 231–236 (2003).
Sanfilippo, A. & Mazzoleni, R. Giorn. Microbiol. 12, 83 (1964).
Parisi, B. & Soller A. Giorn. Microbiol. 12, 183–194 (1964).
Morita, J., Tanaka, A., Komano, T. & Oki, T. Agric. Biol. Chem. 43, 2629–2631 (1979).
Weiss, U., Strelitz, F., Flon, H. & Asheshov, I. N. Arch. Biochem. Biophys. 74, 150–157 (1958).
Strelitz, F., Flon, H., Weiss, U. & Asheshov, I. N. J. Bacteriol. 72, 90–94 (1956).
Strelitz, F., Flon, H. & Asheshov, I. N. J. Bacteriol. 69, 280–283 (1955).
Asheshov, I. N., Strelitz, F., Hall, E. & Flon, H. Antibiot. Chemother. 4, 380–394 (1954).
Landsberger, M. et al. Cell 174, 908–916 (2018).
Borges, A. L. et al. Cell, 174, 917–925 (2018).
Erez, A. et al. Nature 541, 488–493 (2017).
 Read the paper: A chemical defence against phage infection
Read the paper: A chemical defence against phage infection
 Phages make a group decision
Phages make a group decision
 Viruses cooperate to defeat bacteria
Viruses cooperate to defeat bacteria