- NEWS AND VIEWS
Location matters for insulin-producing cells
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 564, 50-51 (2018)
doi: https://doi.org/10.1038/d41586-018-07490-y
References
Hogan, B. L. M. Cell 96, 225–233 (1999).
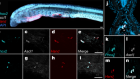
Mamidi, A. et al. Nature https://doi.org/10.1038/s41586-018-0762-2 (2018).
Shih, H. P., Wang, A. & Sander, M. Annu. Rev. Cell Dev. Biol. 29, 81–105 (2013).
Pan, F. C. & Wright, C. Dev. Dyn. 240, 530–565 (2011).
Zhou, Q. et al. Dev. Cell 13, 103–114 (2007).
Totaro, A., Panciera, T. & Piccolo, S. Nature Cell Biol. 20, 888–899 (2018).
Huveneers, S. & Danen, E. H. J. J. Cell Sci. 122, 1059–1069 (2009).
Sneddon, J. B. et al. Cell Stem Cell 22, 810–823 (2018).
 Read the paper: Mechanosignalling via integrins directs fate decisions of pancreatic progenitors
Read the paper: Mechanosignalling via integrins directs fate decisions of pancreatic progenitors
 Peptide secretion triggers diabetes
Peptide secretion triggers diabetes
 The healthy diabetic cavefish conundrum
The healthy diabetic cavefish conundrum