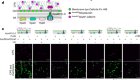
- NEWS AND VIEWS
Tissue ‘melting’ sculpts embryo
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 561, 315-316 (2018)
doi: https://doi.org/10.1038/d41586-018-06108-7
References
Park, J.-A. et al. Nature Mater. 14, 1040–1048 (2015)
Mongera, A. et al. Nature 561, 401–405 (2018).
Steventon, B. et al. Development 143, 1732–1741 (2016).
Tada, M. & Heisenberg, C. P. Development 139, 3897–3904 (2012).
Collinet, C. et al. Nature Cell Biol. 17, 1247–1258 (2015).
Lye, C. M. et al. PLoS Biol. 13, e1002292 (2015).
Bénazéraf, B. et al. Nature 466, 248–252 (2010).
Jülich, D. et al. Dev. Cell 34, 33–44 (2015).
Crest, J. et al. eLife 6, e24958 (2017).
Clément, R. et al. Curr. Biol. 27, 3132–3142.e4 (2017).
Cohen-Addad, S., Höhler, R. & Pitois, O. Annu. Rev. Fluid Mech. 45, 241–267 (2013).
van Hecke, M. J. Phys. Condens. Matter 22, 033101 (2010).
Bi, D., Lopez, J. H., Schwarz, J. M. & Manning, M. L. Nature Phys. 11, 1074–1079 (2015).
Farhadifar, R., Röper, J.-C., Aigouy, B., Eaton, S. & Jülicher, F. Curr. Biol. 17, 2095–2104 (2007).
 Read the paper: A fluid-to-solid jamming transition underlies vertebrate body axis elongation
Read the paper: A fluid-to-solid jamming transition underlies vertebrate body axis elongation
 A gut feeling for cellular fate
A gut feeling for cellular fate
 Mechanics drives cell differentiation
Mechanics drives cell differentiation