- NEWS AND VIEWS
Lessons for cancer drug treatment from tackling a non-cancerous overgrowth syndrome
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 558, 523-525 (2018)
doi: https://doi.org/10.1038/d41586-018-05365-w
References
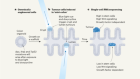
Venot, Q. et al. Nature 558, 540–546 (2018).
Bianconi, E. et al. Ann. Hum. Biol. 40, 463–471 (2013).
Samuels, Y. et al. Science 304, 554 (2004).
Rozengurt, E., Soares, H. P. & Sinnet-Smith, J. Mol. Cancer Ther. 13, 2477–2488 (2014).
Kurek, K. C. et al. Am. J. Hum. Genet. 90, 1108–1115 (2012).
Lindhurst, M. J. et al. Nature Genet. 44, 928–933 (2012).
Rivière, J.-B. et al. Nature Genet. 44, 934–940 (2012).
Keppler-Noreuil, K. M. et al. Am. J. Med. Genet. A 167, 287–295 (2015).
De Santis, M. C. et al. Cancers 9, 30 (2016).
Castillo, S. D. et al. Sci. Transl. Med. 8, 332ra43 (2016).
Gerstung, M. et al. Preprint at bioRxiv https://doi.org/10.1101/161562 (2017).
Jamal-Hanjani, M. et al. N. Engl. J. Med. 376, 2109–2121 (2017).
Yehia, L. & Eng, C. Endocr. Relat. Cancer https://doi.org/10.1530/ERC-18-0162 (2018).
Sansregret, L., Vanhaesebroeck, B. & Swanton, C. Nature Rev. Clin. Oncol. 15, 139–150 (2018).
Ortega-Molina, A. et al. Cell Metab. 21, 558–570 (2015).
Bettedi, L. & Foukas, L. C. Biogerontology 18, 913–929 (2017).
Competing Interests
Bart Vanhaesebroeck is a consultant to Karus Therapeutics (Oxford, UK), iOnctura (Geneva, Switzerland) and Venthera (Palo Alto, California).
 Read the paper: Targeted therapy in patients with PIK3CA-related overgrowth syndrome
Read the paper: Targeted therapy in patients with PIK3CA-related overgrowth syndrome
 When restriction is good
When restriction is good
 Mapping the path to a longer life
Mapping the path to a longer life