- NEWS AND VIEWS
Yeast chromosome numbers minimized using genome editing
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 560, 317-318 (2018)
doi: https://doi.org/10.1038/d41586-018-05309-4
References
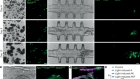
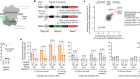
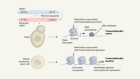
Shao, Y. et al. Nature 560, 331–335 (2018).
Luo, J., Sun, X., Cormack, B. P. & Boeke, J. D. Nature 560, 392–396 (2018).
Goffeau, A. et al. Science 274, 546–567 (1996).
Pryde, F. E. & Louis, E. J. EMBO J. 18, 2538–2550 (1999).
Naseeb, S. et al. Mol. Biol. Evol. 33, 1679–1696 (2016).
Duan, Z. et al. Nature 465, 363–367 (2010).
Greig, D. Heredity 102, 39–44 (2009).
Yue, J.-X. et al. Nature Genet. 49, 913–924 (2017).
Liti, G., Barton, D. B. & Louis, E. J. Genetics 174, 839–850 (2006).
Gordon, J. L., Byrne, K. P. & Wolfe, K. H. PLoS Genet. 7, e1002190 (2011).
 Read the paper: Creating a functional single-chromosome yeast
Read the paper: Creating a functional single-chromosome yeast
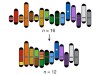 Read the paper: Karyotype engineering by chromosome fusion leads to reproductive isolation in yeast
Read the paper: Karyotype engineering by chromosome fusion leads to reproductive isolation in yeast
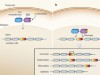 Synthetic yeast genome reveals its versatility
Synthetic yeast genome reveals its versatility
 Edit the genome to understand it
Edit the genome to understand it