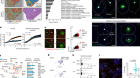
- NEWS AND VIEWS
Tumour tamed by transfer of one T cell
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 558, 193-195 (2018)
doi: https://doi.org/10.1038/d41586-018-05251-5
References
Maus, M. V. & June, C. H. Clin. Cancer Res. 22, 1875–1884 (2016).
Park, J. H. et al. N. Engl. J. Med. 378, 449–459 (2018).
Neelapu, S. S. et al. N. Engl. J. Med. 377, 2531–2544 (2017).
Schuster, S. J. et al. N. Engl. J. Med. 377, 2545–2554 (2017).
Fraietta, J. A. et al. Nature 558, 307–312 (2018).
Hacein-Bey-Abina, S. et al. N. Engl. J. Med. 346, 1185–1193 (2002).
Hacein-Bey-Abina, S. et al. Science 302, 415–419 (2003).
Biasco, L. et al. EMBO Mol. Med. 3, 89–101 (2011).
Scholler, J. et al. Sci. Transl. Med. 4, 132ra53 (2012).
Schröder, A. R. W. et al. Cell 110, 521–529 (2002).
Buscarlet, M. et al. Blood 130, 753–762 (2017).
Eyquem, J. et al. Nature 543, 113–117 (2017).
Ren, J. et al. Clin. Cancer Res. 23, 2255–2266 (2016).
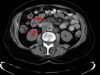 Read the paper: Disruption of TET2 promotes the therapeutic efficacy of CD19-targeted T cells
Read the paper: Disruption of TET2 promotes the therapeutic efficacy of CD19-targeted T cells
 T-cell tweaks to target tumours
T-cell tweaks to target tumours
 Tumour lymph vessels boost immunotherapy
Tumour lymph vessels boost immunotherapy