- NEWS AND VIEWS
Longer life through an odd Pol enzyme
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Rent or buy this article
Prices vary by article type
from$1.95
to$39.95
Prices may be subject to local taxes which are calculated during checkout
Nature 552, 182-183 (2017)
doi: https://doi.org/10.1038/d41586-017-07435-x
References
Warner, J. R. Trends Biochem. Sci. 24, 437–440 (1999).
Filer, D. et al. Nature 552, 263–267 (2017).
McCay, C. M., Crowell, M. F. & Maynard, L. A. J. Nutr. 10, 63–79 (1935).
Kapahi, P. Adv. Exp. Med. Biol. 694, 30–37 (2010).
Mehta, R., Chandler-Brown, D., Ramos, F. J., Shamieh, L. S. & Kaeberlein, M. Adv. Exp. Med. Biol. 694, 14–29 (2010).
Wang, L., Karpac, J. & Jasper, H. J. Exp. Biol. 217, 109–118 (2014).
Tomasetti, C. & Vogelstein, B. Science 347, 78–81 (2015).
Li, H. & Jasper, H. Dis. Model. Mech. 9, 487–499 (2016).
Michels, A. A. Biochem. Soc. Trans. 39, 487–491 (2011).
Rideout, E. J., Marshall, L. & Grewal, S. S. Proc. Natl Acad. Sci. USA 109, 1139–1144 (2012).
Khanna, A., Johnson, D. L. & Curran, S. P. Cell Rep. 9, 2180–2191 (2014).
Cai, Y. & Wei, Y.-H. Oncotarget 7, 10812–10826 (2016).
Cai, Y. & Wei, Y.-H. Aging 7, 133–143 (2015).
Demontis, F., Patel, V. K., Swindell, W. R. & Perrimon, N. Cell Rep. 7, 1481–1494 (2014).
Tiku, V. et al. Nature Commun. 8, 16083 (2017).
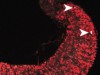 Read the paper: RNA polymerase III limits longevity downstream of TORC1
Read the paper: RNA polymerase III limits longevity downstream of TORC1
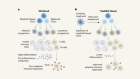
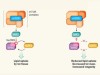 An atypical switch for metabolism and ageing
An atypical switch for metabolism and ageing
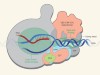 A transcriptional specialist resolved
A transcriptional specialist resolved