
Abstract
T cells are derived from progenitor thymocytes, of which only a minority receive the appropriate TCR signal, undergo positive selection and mature. Owing to the very short lifespan of thymocytes, the prerequisite for positive selection is survival. TCR signal-induced Bcl-2 expression is believed to play a dominant role in the survival of positively selecting thymocytes, but how Bcl-2 is directly regulated is unknown. Here we report that the immediate early gene (IEG) c-Fos can stimulate the expression of Bcl-2, depending on a specific AP-1-binding site in the Bcl-2 promoter. In c-Fos transgenic (Fos-Tg) mice, c-Fos binds to this site and promotes the expression of Bcl-2. As a result, Fos-Tg thymocytes exhibited enhanced survival, and more mature single-positive (SP) thymocytes were generated, even on a unique TCR background. The TCR repertoire remained normal in Fos-Tg mice. Our results identified c-Fos as the mediator of the stimulatory effect of TCR signaling on Bcl-2 expression. Therefore, c-Fos, as an IEG, because of its early response ability, can quickly rescue the survival of short-lived thymocytes during positive selection. Our results provide novel insight into the mechanism regulating the survival of positively selecting thymocytes.
Similar content being viewed by others

Introduction
T cells are generated from progenitor thymocytes during development, with TCR recombination and selection occurring in the thymus 1, 2. Early CD4−CD8− double-negative (DN) thymocytes rearrange the T cell receptor β (Tcrb) gene 3, proliferate and differentiate into CD4+CD8+ double-positive (DP) cells, which account for a majority of the thymocytes. DP thymocytes expressing the αβTCR undergo selection to generate a functional T-cell repertoire. Thymocyte selection is controlled by TCR signals that are induced by the interaction between the TCR and complexes of self-peptide and the major histocompatibility complexes (pMHC). Signals that are too weak do not support the survival of thymocytes, and these cells will die because of neglect. Strong signals through the TCR induce negative selection so that self-reactive thymocytes are eliminated. Only a minority of thymocytes receive the appropriate TCR signal, survive and mature into CD4 or CD8 single-positive (SP) T cells. This process is called positive selection. During positive selection, one of the most important events is survival. It has been established that the anti-apoptotic molecule Bcl-2 is responsible for survival during positive selection 4, 5. Most DP thymocytes express a very low level of Bcl-2, but after receiving a TCR signal DP thymocytes increase their expression of Bcl-2. Bcl-2 ensures the survival of positively selected thymocytes, but it is not involved in thymocyte differentiation and cannot overcome negative selection 6. Although the essential role of Bcl-2 in thymocyte survival has been identified, how TCR signals induce the expression of Bcl-2 in thymocytes is still obscure.
The immediate early gene (IEG) c-Fos encodes a transcriptional factor. c-Fos protein can form the AP-1 heterodimer with Jun proteins and stimulate the expression of many genes during TCR stimulation-mediated T-cell responses 7, 8, which is a very quick process 9. Just like Bcl-2, c-Fos is upregulated by TCR signals in positively selecting DP thymocytes 10, 11. Although the purpose of this upregulation is unknown, it has been reported that c-Fos plays an essential role in protecting DP thymocytes from Ca2+ and cAMP-induced apoptosis but not from TCR-mediated death mimicking negative selection 12. The spontaneous and induced death of Fos−/− DP thymocytes is also increased. In addition, Fos−/− mice display a reduction in thymus cellularity, and the frequency of DP thymocytes is decreased by 90% in these mice 13. However, how c-Fos protects thymocytes from apoptosis is still unknown.
In this report, we found that c-Fos could directly promote the expression of Bcl-2 by binding to an AP-1-binding site in the Bcl-2 promoter. In c-Fos transgenic (Fos-Tg) mice, Bcl-2 expression was elevated, survival of thymocytes was improved and, in turn, significantly more cells were positively selected. Meanwhile, Vβ staining revealed that the TCR repertoire was not changed in Fos-Tg mice.
Results
c-Fos activates the promoter of Bcl-2
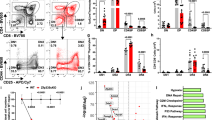
As both Bcl-2 and c-Fos were upregulated in positively selecting DP thymocytes and c-Fos can protect DP thymocytes from apoptosis, we hypothesized that c-Fos might regulate the expression of Bcl-2 during positive selection. The expression profiles of c-Fos and Bcl-2 at various stages of thymocyte development were analyzed. Flow-cytometry assays revealed that c-Fos and Bcl-2 expression levels changed in a similar manner; both were at very low levels in the DP stage, at mild levels in positively selecting CD4lo8+ and CD4+8lointermediates and at high levels in the DN stage (Figure 1A). The covariation relationship between the expression profiles of c-Fos and Bcl-2 further led us to predict that c-Fos may regulate the expression of Bcl-2. To verify this, we constructed a luciferase reporter controlled by the Bcl-2 promoter. Overexpression of c-Fos in HeLa cells led to a four-fold increase in the activation of the Bcl-2 promoter (Figure 1B). Moreover, such an effect was dose dependent. As the levels of c-Fos increased, Bcl-2 promoter activity was also elevated. This result revealed that c-Fos could activate the Bcl-2 promoter. As c-Fos itself cannot form homodimers and bind DNA, it may function together with Jun proteins in HeLa cells.

c-Fos upregulated Bcl-2 expression. (A) Thymocytes from B6 mice were stained for CD4, CD8 and intracellular c-Fos, Bcl-2 or the corresponding isotype control. The histograms display the expression of c-Fos (top panel) and Bcl-2 (bottom panel) in different thymocyte subsets (including DP, CD4lo8+, CD4+8lo and DN). (B) HeLa cells were transfected with the pGL3 vector or the pGL3 construct containing the Bcl-2 promoter sequence and increasing amounts (wedge) of the c-Fos expression construct. Reporter activity was measured 24 h after transfection and normalized to Renilla luciferase activity. Data represent mean values ± SEM of three independent experiments.
Identification of the c-Fos-binding site in the Bcl-2 promoter
To test whether c-Fos directly regulates Bcl-2 expression, the promoter sequence of Bcl-2 from –3 000 to +1 000 was analyzed using computational software. Three potential AP-1 binding sites — S1, S2 and S3 — were found (Figure 2A). Bcl-2 promoter mutants were constructed according to these binding sites and were then analyzed with luciferase assays (Figure 2A). Deletion of either S1 or S1 and S2 did not impair the stimulatory effect of c-Fos overexpression on Bcl-2 promoter activity. However, mutation of S3 resulted in the complete loss of Bcl-2 inducibility by c-Fos. Moreover, in the absence of transfected c-Fos, the baseline activity of the S3 mutant promoter was much lower than that of the wild-type Bcl-2 promoter. This may be because of the high endogenous c-Fos expression in HeLa cells. Indeed, the level of endogenous c-Fos in HeLa cells was relatively high and was easily detected by western blotting (data not shown). These results suggest that c-Fos directly regulates the expression of Bcl-2 and that this regulation is dependent on the S3 AP-1-binding site.

Identification of the c-Fos-binding site in the Bcl-2 promoter. (A) Top, a diagram of the potential AP-1-binding sites in the Bcl-2 promoter. Bottom, HeLa cells transfected with reporter constructs that contained the wild-type Bcl-2 gene promoter sequence (pGl3-Bcl-2) or three different deletions or mutations of the AP-1 sites of the Bcl-2 promoter, as well as the expression vector (pcDNA) or expression constructs for c-Fos (pcDNA-c-Fos). The mutant forms of the Bcl-2 promoter are indicated in the left panel. Reporter gene activity was measured 24 h after transfection and normalized to Renilla luciferase activity. Data represent mean values ± SEM of three to five independent experiments. (B) Chromatin-immunoprecipitation assays with B6 mouse CD69− and CD69hi DP thymocytes. Samples were obtained with anti-c-Fos antibody or a normal IgG control. The amounts of S1 and S3 were estimated with real-time PCR. Data are presented as 'fold increase' (mean values ± SEM from two experiments) relative to the input.
After identifying the AP-1-binding site in the Bcl-2 promoter with luciferase assays, we tested whether c-Fos bound to this site in vivo in positively selecting thymocytes. CD69hi DP thymocytes (representing DP thymocytes that had received a TCR signal) were sorted and used in IgG- or c-Fos antibody-mediated chromatin-immunoprecipitation (ChIP) assay. Substantial enrichment of the S3 segment by the c-Fos antibody was detected as compared with the IgG control (Figure 2B). As the amount of c-Fos in non-signaled CD69− DP cells was very low, there was no obvious binding of c-Fos to the Bcl-2 promoter in these cells (Figure 2B). As a negative control, no enrichment of the S1 segment was detected, which was consistent with the result from the luciferase assays. The above data further support the fact that c-Fos could mediate the stimulatory effect of the TCR signal on the expression of Bcl-2.
Ectopic c-Fos expression elevates Bcl-2 expression in thymocytes
As c-Fos could directly activate the Bcl-2 promoter in vitro, we determined whether such a function of c-Fos existed in vivo. Fos-Tg mice in which ectopic c-Fos expression was under the control of the human CD2 promoter and was restricted to only T lineage cells were created. The c-Fos protein level in transgenic thymocytes was comparable to that in wild-type thymocytes stimulated with phorbol ester (PMA) and ionomycin (IM) for 2 h; therefore, ectopic c-Fos expression mimicked physiological conditions (Figure 3A). Meanwhile, ChIP assay of DP thymocytes showed that more c-Fos proteins bound to the Bcl-2 promoter in Fos-Tg mice (Figure 3B).

The expression of Bcl-2 was enhanced in thymocytes from Fos-Tg mice. (A) Fresh thymocytes from Fos-Tg and littermate controls or thymocytes stimulated with PMA/IM for different times were analyzed by immunoblotting for expression of c-Fos. Actin served as a loading control. (B) Chromatin immunoprecipitation assays with Fos-Tg or littermate control DP thymocytes for c-Fos antibody-mediated S3 enrichment. Data are presented as 'fold increase' (mean values ± SEM from three experiments) relative to the input. (C) Thymocytes from Fos-Tg and littermate mice were stained for CD4, CD8 and intracellular Bcl-2 or an isotype control. The histogram displays the expression of Bcl-2 in different thymocyte subsets, as indicated. (D) Thymocytes from Fos-Tg and littermate mice were stained for PI and Annexin V. Gated on PI negative cells, the staining of Annexin V in the different thymocyte subsets is shown in the histogram. The percentage of Annexin V-positive thymocytes is indicated. Litt, littermate; Fos-Tg, c-Fos transgenic.
Bcl-2 expression in Fos-Tg and littermate thymocytes was analyzed by flow cytometry. Results showed that Bcl-2 was upregulated in Fos-Tg mice throughout thymocyte positive selection, including DP, CD4lo8+ and CD4+8lo thymocytes (Figure 3C). Accordingly, Annexin V staining revealed that the survival of Fos-Tg thymocytes was consequently improved (Figure 3D). Interestingly, the CD4loCD8+ thymocyte subpopulation was affected to a greater extent than other populations in the Fos-Tg mice. This may be because the baseline expression of c-Fos and Bcl-2 in CD4loCD8+ thymocytes is lower than that in CD4+CD8lo thymocytes, as shown in Figure 1A. After c-Fos is overexpressed, Bcl-2 is upregulated to a saturating level and the amount of Bcl-2 expression is comparable between these two subpopulations (Figure 3C). As a result, the enhancement of Bcl-2 expression and cell survival was more obvious in CD4loCD8+ thymocytes in Fos-Tg mice. In fact, survival of the CD4loCD8+ subpopulation was also more obviously enhanced than other subpopulations 14 in Bcl-2 transgenic mice.
Thymocyte positive selection is promoted in Fos-Tg mice
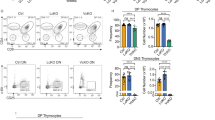
To verify the effect of increased Bcl-2 levels on thymocyte development, the expression of CD4 and CD8 in thymocytes from Fos-Tg and littermate mice was examined. The percentage of both CD4SP and CD8SP thymocytes was increased substantially in Fos-Tg mice, from 4.9% to 16.3% and from 1.51% to 17.5%, respectively. Moreover, the positive selection intermediates also increased (Figure 4A). The absolute number of CD4SP and CD8SP cells increased dramatically (Figure 4B). Fos-Tg SP thymocytes were TCR high and HSA low, indicating that these SP cells were mature (Figure 4C and 4D). Consistently, the number of peripheral lymphocytes also increased slightly (data not shown). We also analyzed positive selection markers in Fos-Tg mice and found that CD69 and CD25 were obviously upregulated in Fos-Tg mice (Figure 4E). Therefore, increased Bcl-2 expression promoted thymocyte positive selection and gave rise to a larger number of mature SP thymocytes in Fos-Tg mice. In these assays, we also found that c-Fos overexpression had a more profound effect on the CD8 lineage; the extent of increase of CD8SP cells was more than that of CD4SP cells. These phenotypes of the thymocytes from Fos-Tg mice were similar to those in Bcl-2 transgenic mice, further supporting the fact that c-Fos enhances the survival of positively selecting thymocytes by upregulating Bcl-2.

Fos-Tg mice increased the generation of mature SP thymocytes. (A) Thymocytes from Fos-Tg and littermate mice were stained for CD4 and CD8. The dot plot displays the expression of CD4 and CD8. The percentage of gated cells that fell into each quadrant is indicated. (B) Absolute numbers of total, CD4SP and CD8SP thymocytes from Fos-Tg and littermate mice are shown. Data are presented as mean values ± SEM from four groups of mice. *P < 0.01, versus the littermate control. (C) The histogram displays the expression of TCRβ on the surface of CD4SP and CD8SP thymocytes from Fos-Tg and littermate mice. (D) The histogram displays the expression of HSA on the surface of CD4SP and CD8SP thymocytes from Fos-Tg and littermate mice. (E) Thymocytes from Fos-Tg and littermate mice were analyzed by flow cytometry for the expression of CD69 and CD25. Litt, littermate; Fos-Tg, c-Fos transgenic.
The TCR repertoire is normal in Fos-Tg mice
As thymocyte selection is regulated by the avidity of the TCR-MHC/peptide interaction, a TCR repertoire shift may also affect positive selection. We thus investigated the TCR repertoire in Fos-Tg and littermate mice using Vβ-specific antibodies. Seven Vβ-specific antibodies were used to stain the thymocytes and lymphocytes from Fos-Tg and littermate mice. However, no difference was found in the TCR repertoire (Figure 5A and 5B). Therefore, ectopic expression of c-Fos did not change the TCR repertoire.

The TCR repertoire was normal in Fos-Tg mice. (A) Thymocytes from Fos-Tg and littermate mice were stained for CD3, PI and specific Vβs. PI negative and CD3+ thymocytes were gated, and the percentages of specific Vβ-positive thymocytes are shown. Data are presented as mean values ± SEM from two groups of mice. (B) T lymphocytes from Fos-Tg and littermate mice lymph nodes were stained for CD3, PI and specific Vβs. PI negative and CD3+ T lymphocytes were gated, and the percentages of specific Vβ-positive lymphocytes are shown. Data are presented as mean values ± SEM from two groups of mice. Litt, littermate; Fos-Tg, c-Fos transgenic.
Generation of SP thymocytes is promoted by c-Fos in a TCR transgenic background
Our earlier data showed that c-Fos could enhance Tcrb recombination, and in turn promote the development of DN thymocytes, which may also indirectly contribute to the increased number of SP thymocytes 15. To elucidate this possibility, Fos-Tg mice were crossed to OT-1 TCR transgenic mice. Under this TCR transgenic background, the endogenous Tcrb recombination was prohibited and thus c-Fos could not function in this process. However, expression of the c-Fos transgene still gave rise to an increased number of mature SP thymocytes (Figure 6A and 6B). This result indicates that c-Fos mediates the stimulatory effect of the TCR signal on Bcl-2 expression and thymocyte positive selection independent of its role in Tcrb recombination.

Ectopic expression of c-Fos enhanced the generation of mature SP thymocytes. (A) Thymocytes from Fos-Tg and littermate mice on the OT-1 TCR transgenic background were stained for CD4 and CD8. The dot plot displays the expression of CD4 and CD8. The percentage of gated cells that fell into each quadrant is given. (B) Absolute numbers of total and CD8SP thymocytes from littermate and Fos-Tg mice on the OT-1 TCR transgenic background are shown. Data are presented as mean values ± SEM from three groups of mice. *P < 0.01, versus the littermate control. Litt, littermate; Fos-Tg, c-Fos transgenic.
Discussion
During the positive selection of thymocytes, DP thymocytes that have received a TCR signal upregulate the expression of Bcl-2 to survive and mature to SP thymocytes. Survival is one of the necessary and most important events in positive selection. However, which molecule mediates the TCR signal and directly regulates the expression of Bcl-2 during this process is unknown. Our study approached this longstanding question and identified c-Fos as the molecule. c-Fos could bind to the promoter of Bcl-2 and activate its expression. Results from Fos-Tg mice verified this role of c-Fos, and more mature SP thymocytes were generated as a result of the improved survival of thymocytes.
Several IEGs, including Nur77 and Egr1, have been reported to function in thymocyte development. IEGs, because of their early response ability, quickly regulate thymocyte selection after cells receive a TCR signal. Egr1 alters the threshold of signaling required for positive selection 16. It has also been reported that Egr1 may regulate Bcl-2 expression, but direct evidence is lacking 17. The orphan nuclear receptor Nur77 is involved in thymocyte negative selection 18, 19. Although the IEG c-Fos has been found to be expressed differentially during thymocyte selection and to protect DP thymocytes from apoptosis, its function in thymocyte selection and the underlying mechanism are unknown. Here, we identified c-Fos as a direct regulator of DP thymocytes that have received a signal during positive selection. Such a role for c-Fos is of special importance for thymocyte development. As DP thymocytes have a very short lifespan and can only survive for 2-3 days if they do not receive a TCR signal 20, c-Fos must quickly upregulate the expression of Bcl-2 to allow survival for further maturation. As an IEG, c-Fos fits this description.
The role of c-Fos in promoting cell survival is cell type-specific. It has been reported that c-Fos-induced apoptosis in several non-lymphoid transformed cell systems through unknown mechanisms 21, 22, and transgenic c-Fos can even induce apoptosis in germinal center B cells 23. However, c-Fos could promote neuronal survival 24. Our study here shows that c-Fos selectively promotes the survival of thymocytes during positive selection.
Materials and Methods
Mice
OT-1 TCR transgenic mice were initially obtained from the Jackson Laboratory and were subsequently maintained by breeding with our own colony. Fos-Tg mice were generated by the overexpression of c-Fos under the control of the human CD2 promoter and locus control region in the transgenic expression vector p29Δ2 (gift of Paul Love). Except where otherwise indicated, mice were analyzed between 5 and 10 weeks of age and were heterozygous for the transgene(s) they carried. All mice were maintained in SPF facilities and were genotyped before use. All animal experiments were approved by the institutional animal use committee of the Shanghai Institutes for Biological Sciences (Chinese Academy of Sciences).
Sequence analysis
Potential AP-1-binding sites were identified using the TFSSEARCH System (http://mbs.cbrc.jp/research/db/TFSEARCH.html) and by alignment with the AP-1-binding consensus sequences.
Plasmids
The cDNA encoding c-Fos was generated by RT-PCR of thymocyte RNA preparations and cloned into the mammalian expression vector pcDNA3.0. The promoter sequence of Bcl−2 from −2 166 to +765 was cloned into pGL3-basic to construct pGL3-Bcl−2. The deletion forms were generated as indicated in the figures. The S3 AP-1-binding site mutation was from 5′-TGACTAA-3′ to 5′-AGACCAA-3′.
Cell culture, transfections and luciferase assays
HeLa cells were maintained in Dulbecco's modified Eagle's medium and 10% fetal calf serum. Cells were transfected with Lipofectamine2000 (Invitrogen) according to the manufacturer's instructions. Luciferase activity was measured 24 h after transfection with the Dual Luciferase Assay Kit (Promega) and a luminometer (Turner Design).
ChIP assay
ChIP assays were carried out, with modification, according to the manufacturer's instructions (Upstate Biology). 2 × 105 purified thymocytes were crosslinked with 1% formaldehyde on ice for 30 min and resuspended in 200 μl of ChIP lysis buffer. The lysates were sonicated on ice to obtain DNA fragments of approximately 500 bp and were immunoprecipitated with IgG or anti-c-Fos antibody. The recovered DNA was detected by quantitative real-time PCR. For real-time PCR, the data were normalized to those obtained with the corresponding DNA input control. The primer pairs were designed to cover approximately ± 100 bp of S1 and S3. Primers used were as follows (site, forward primer/reverse primer): S1, 5′-TAA AGG TGG AAG GGG AGA ATG G-3′/5′-AGG CTT GCT GGA TTA CTG TGG G-3′; S3, 5′-CTG GGT GCT CAT GTG GTA TCT G-3′/5′-TGA TGT CCA CAA CCA GCA AGG-3′.
Antibodies
The following monoclonal antibodies were used for staining: anti-CD4 (RM4-5), anti-CD8a (53–6.7), anti–TCRβ (H57-597), anti-Vβ4TCR (CTVB4), anti-Vβ5.1/5.2 TCR (MR9-4), anti-Vβ6 TCR (RR4-7), anti-Vβ7 TCR (TR310), anti-Vβ8.1/8.2 TCR (KJ16), anti-Vβ12 TCR (CTVB12b), anti-Vβ14 TCR (14-2), anti-HSA (M1/69), anti-Bcl-2 (3F11) and hamster IgG isotype control; all antibodies were from BD Pharmingen. Antibodies to c-Fos (4) and normal rabbit IgG were from Santa Cruz.
Thymocyte preparation, purification and staining
Single-cell thymocyte or splenocyte suspensions were prepared and stained as described 25. Cell fluorescence was typically acquired on 1 × 105 cells on a two-laser FACSCalibur (BD Biosciences) with four-decade logarithmic amplification. Data were analyzed with FlowJo software. Live cells were identified by forward light scatter and propidium iodide gating. CD69− and CD69hi DP thymocytes were purified by cell sorting with a FACSAria (BD Biosciences). Cell purity, as assessed by surface staining and flow cytometry, was greater than 90%.
Statistics
Student's t-test was used for the comparison of two independent groups. For all tests, a P value of <0.05 was considered statistically significant.
References
Starr TK, Jameson SC, Hogquist KA . Positive and negative selection of T cells. Annu Rev Immunol 2003; 21:137–176.
Goldrath AW, Bevan MJ . Selecting and maintaining a diverse T-cell repertoire. Nature 1999; 402:255–262.
Schlissel MS . Regulating antigen-receptor gene assembly. Nat Rev Immunol 2003; 3:890–899.
Linette GP, Grusby MJ, Hedrick TH, et al. Bcl-2 is upregulated at the CD4+ CD8+ stage during positive selection and promotes thymocyte differentiation at several control points. Immunity 1994; 1:197–205.
Williams O, Norton T, Halligey M, Kioussis D, Brady HJ . The action of Bax and Bcl-2 on T cell selection. J Exp Med 1998; 188:1125–1133.
Sentman CL, Shutter JR, Hockenbery D, Kanagawa O, Korsmeyer SJ . Bcl-2 inhibits multiple forms of apoptosis but not negative selection in thymocytes. Cell 1991; 67:879–888.
Chiu R, Boyle WJ, Meek J, et al. The c-fos protein interacts with c-Jun/AP-1 to stimulate transcription of AP-1 responsive genes. Cell 1988; 54:541–552.
Moore JP, Menzel GE, Hesketh TR, Metcalfe JC . c-fos gene activation in murine thymocytes by a mechanism independent of protein kinase C or a Ca2+ signal. FEBS Lett 1988; 233:64–68.
Jain J, Valge-Archer VE, Rao A . Analysis of the AP-1 sites in the IL2 promoter. J Immunol 1992; 148:1240–1250.
Nunomura S, Sato T, Habu S . Molecular basis for functional maturation of thymocytes: increase in c-fos translation with positive selection. J Immunol 2000; 164:5590–5595.
Rincon M, Flavell R . Regulation of AP-1 and NFAT transcription factors during thymic selection of T cells. Mol Cell Biol 1996; 16:1074–1084.
Ivanov VN, Nikolić-Zugić J . Transcription factor activation during signal-induced apoptosis of immature CD4+CD8+ thymocytes. J Biol Chem 1997; 272:8558–8566.
Wang ZQ, Ovitt C, Grigoriadis AE, et al. Bone and haematopoietic defects in mice lacking c-fos. Nature 1992; 360:741–745.
Liu X, Bosselut R . Duration of TCR signaling controls CD4-CD8 lineage differentiation in vivo. Nat Immunol 2004; 2:280–288.
Wang X, Xiao G, Zhang Y, et al. Regulation of Tcrb recombination ordering by c-Fos dependent RAG deposition. Nat Immunol 2008; 9:794–801.
Miyazaki T, Lemonnier FA . Modulation of thymic selection by expression of an immediate-early gene, early growth response 1(Egr-1). J Exp Med 1998; 188:715–723.
Bettini M, Xi H, Milbrandt J, Kersh GJ . Thymocytes development in early growth reponse gene 1-deficient mice. J Immunol 2002; 169:1713–1720.
Liu ZG, Smith SW, McLaughlin KA, Schwartz LM, Osborne BA . Apoptotic signals delivered through the T-cell receptor of a T-cell hybrid require the immediate-early gene nur77. Nature 1994; 367:281–284.
Amsen D, Calvo CR, Osborne BA, Kruisbeek AM . Costimulatory signals are required for induction of transcription factor Nur77 during negative selection of CD4+CD8+ thymocytes. Proc Natl Acad Sci USA 1999; 96:622–627.
von Boehmer H, Teh HS, Kisielow P . The thymus selects the useful, neglects the useless and destroys the harmful. Immunol Today 1989; 10:57–61.
Smeyne RJ, Vendrell M, Hayward M, et al. Continuous c-fos expression precedes programmed cell death in vivo. Nature 1993; 363:166–169.
Preston GA, Lyon TT, Lang JE, et al. Induction of apoptosis by c-Fos protein. Mol Cell Biol 1996; 16:211–218.
Inada K, Okada S, Phuchareon J, et al. c-Fos induces apoptosis in germinal center B cells. J Immunol 1998; 161:3853–3861.
Zhang J, Zhang D, McQuade JS, et al. c-fos regulates neuronal excitability and survival. Nat Genet 2002; 30:416–420.
Liu X, Adams A, Wildt KF, et al. Restricting Zap70 expression to CD4+ CD8+thymocytes reveals a T cell receptor-dependent proofreading mechanism controlling the completion of positive selection. J Exp Med 2003; 197:363–373.
Acknowledgements
We thank our colleagures, Q Yuan for animal husbandry and Z Tan for cell sorting. This research was supported in part by the National Natural Science Foundation of China (30671913, 30623003), the Ministry of Science and Technology (2006CB504303, 2007CB815802), the National High-Tech Research and Development Program of China (2007AA02Z167) and the Chinese Academy of Sciences ('The Hundred Talents Program', 180KSCX2-YW-R-108).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wang, X., Zhang, Y., Xiao, G. et al. c-Fos enhances the survival of thymocytes during positive selection by upregulating Bcl-2. Cell Res 19, 340–347 (2009). https://doi.org/10.1038/cr.2008.322
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cr.2008.322
