
Abstract
The effects of antisense oligonucleotide to insulin-like growth factor II (IGFII) to induce apoptosis in human ovarian cancer cells were evaluated. Antiproliferation effects of antisense to IGFII in ovarian cancer AO cells were determined by 3H-thymidine incorporation. Apoptosis of the IGFII antisense-treated cells was quantitated by both nuclear condensation and flow cytometry after cells were stained with propidium iodide. IGFII antisense (4.5 μM) treatment of 48 h maximally inhibited proliferation of AO cells. More than 25% of IGFII antisense-treated cells (4.5 μM for 24 h) had undergone apoptosis, whereas less than 3% of the cells were apoptotic in either IGFII sense-treated cells or untreated cells. Antisense oligonucleotide to IGFII significantly inhibited cell proliferation and induced apoptosis in human ovarian cancer AO cell. These data suggest that IGFII may be a potential target in treatment of ovarian cancer and antisense oligonucleotide to IGFII may serve as a therapeutic approach.
Similar content being viewed by others
Introduction
Ovarian cancer is one of the most common fatal gynesologic malignancies in the world1. Clinical treatment of ovarian cancer, though with great progress, has been impeded by resistence to chemical therapy. Recently, induction of apoptosis in cancer cells has become not only a hot topic in basic research but also a focal point in the clinical treatment. Antisense approach has been recently adopted to induce apoptosis in some cancer cells, showing the promosing future for its clinical application.
IGFII is an important growth and/or differentiation factor during normal fetal development and also plays essential roles in many primary human malignant tumors as well as in ovarian cancer cells2, 3. It is thus of great interest to assess the potential effects of inhibition of IGFII expression by antisense oligonucleotide on proliferation and apoptosis in human ovarian cancer cells.
Materials and Methods
Cell culture and drug treatment
Human ovarian cancer cell line, AO, was obtained from the Cell Bank at the Chinese Academy of Sciences. AO cell line was cultured in RPMI 1640 (Gibco) supplemented with 10% heat-inactivated fetal calf serum (Evergreen), 100 units/ ml penicillin, 100 μg/ml streptomycin, and 2 mM glutamine. Exponentially growing AO cells (2 x 104 cells/ml) were changed into the medium with 2% heat-inactivated fetal calf serum and were treated with different concentrations of either IGFII antisense or IGFII sense oligodeoxynuceotides after 12 h.
Sequences of S-oligonucleotides
The sequences of S-oligonucleotides, synthesized by Sangon Ltd Canada,were as follows:
IGFII antisense: 3' TAC CCT TAC/G GGT TAC CCC TTC 5'
IGFII sense: 5' ATG GGA ATG/C CCA ATG GGG AAG 3'
Assay for inhibition of 3H-thymidine incorporation
AO cells were plated in sextuplicate wells in 96-well microtest plates, and treated as described4. At various intervals after either IGFII antisense or IGFII sense were added, the cells were pulsed with 2μCi [3H]-thymidine/well (22 Ci/mmol; Shanghai Institute of Nuclear Sciences, Chinese Academy of Sciences), trypsinized, and harvested on strips of fiberglass filter paper with a multiple automated sample harvesters. The radioactivity of each sample was measured in a liquid scintillation counter.
Morphological analysis of apoptotic cells
Random fields of control or treated cells were photographed through a 40 x objective lens in both phase and fluoresent modes. Apoptotic cells, after staining with propidium iodide (PI, Sigma) as described5, were much smaller than the viable cells and presented a condensed chromatin and the fragmented nuclear chromatin bodies6.
Quantitative analysis of apoptosis
The cells (2 x 106) treated with 4.5 μM IGFII antisense or 4.5 μM IGFII sense or none were washed twice with PBS containing 0.1% glucose, and then fixed in 1 ml of ice-cold ethanol overnight at 4 °C. The fixed cells were pelleted and resuspended in 0.5 ml of PBS containing 0.1% glucose, 30 μg/ml PI, and 1 mg/ml RNAse A (Sigma). The DNA contents of the cells were analyzed by Flow Cytometry (Becton-Dickinson, San Jose, CA) as described7, 8.
Cell viability analysis
Cell viability analysis was performed by conventional trypan blue exclusion9. 3 x 105 cells were plated in 6-well plates. After 24 h, the cells were changed to the medium with 2% heat-inactivated fetal calf serum and incubated for 12 h. IGFII antisense or IGFII sense oligonucleotides were added to the medium and incubated for another 24 h. Then, the cells were trypsinized and viable cells that excluded trypan blue exclude were counted under a phase microscope. Independent experiments were carried out for at least three times, and the data were given as the means ± SEM.
Statistics
Statistical analyses of data were performed using Student's t-test
Results
Inhibition of cell proliferation by IGFII antisense
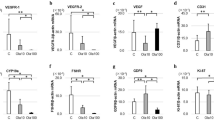
Proliferation of human ovarian cancer AO cells was dramatically inhibited by IGFII antisense treatment (Fig 1) and the cell viability of AO cells was greatly reduced by IGFII antisense treatment too (Fig 2). The inhibition by IGFII antisense was dose- and time-dependent. Significant inhibition occurs at 24 h after application of 1.5 μM IGFII antisense and at 12 h after application of 4.5 μM IGFII antisense. In contrast, treatment of cells with 4.5 μM IGFII sense oligonucleotide for 48 h did not show any significant inhibition. IGFII antisense treatment at 4.5 μM for 24 h (more than 40% inhibition) was used for the subsequent experiments.

Effects of IGFII antisense and IGFII sense on 3H-thymidine incorporation of AO cells. (A) The dose-response curves after 24 h treatment. (B) The time courses of treatments with different concentration of oligonucleotides. The percentages of 3H thymidine incorporation of the treated samples were calculated against untreated cells. At least three independent experiments each done in sextuplicate were performed to obtain means and SEM. Symbols: S, Sense; AS, Antisense.

Effects of IGFII antisense and IGFII sense on cell viability of AO cells. (A) The dose- response curves of effect of IGFII antisense and sense on the cell viability of AO cells after 24 h treatment. (B) The time course of cell viability of AO cells. The percentages of cell viability of the treated samples were calculated against untreated cells. At least three independent experiments each done in sextuplicate were performed to obtain means and SEM. Symbols: S, Sense; AS, Antisense.
Morphological changes induced by IGFII antisense consistent with apoptosis
The effects of IGFII antisense on the morphology of AO cells were examined using fluorescent staining of nuclear DNA with PI. Treatment of AO cells with 4.5 μM IGFII antisense for 24 h (Fig 3) resulted in morphological changes including condensation of chromatin at the nuclear membrane and nuclear fragmentation with groups of isolated pieces of condensed chromatin, which are characteristic of apoptosis

Effects of IGFII antisense and IGFII sense oligonucleotides treatments on the mor- phology of AO cells. Cultured cells were treated for 24 h with 4.5 μM IGFII antisense or 4.5 μM IGFII sense or control medium, fixed with methanol:acetic acid (3:1), and stained for 15 min with PI (30 μg/ml in PBS). The slides loaded with cells were washed, mounted in PBS, and observed under fluorescence microscope. Representa- tive pictures are shown in (A) for control cells, in (B) for 4.5 μM IGFII antisense and in (C) for 4.5 μM IGFII sense-treated cells. The control and 4.5 μM IGFII sense-treated cells show intact nuclei but the 4.5 μM IGFII antisense-treated cells demonstrate signs of apoptosis, with condensation of nuclear masses at the nuclear membrane and nuclear fragmentation.
Flow cytometry analysis of IGFII antisense-induced apoptosis
Flow cytometry was applied to further analyze apoptosis induced by IGFII antisense in human ovarian cancer AO cells. 24 h after incubation of the cells with 4.5 μM IGFII antisense, the apoptotic cells, as represented by the sub-G1 peak in the sample histograms (Fig 4), reached about 25% of the total cell number. The apoptotic cells in the IGFII sense-treated or untreated ovarian cancer AO cells were less than 2% of the total cell population.

Fluorescence histograms of AO cells after treated with 4.5 μM IGFII antisense or 4.5 μM IGFII sense for 24 h. Representative his- tograms of DNA analysis by flow cytometry (n=3) were shown as indicated for con- trol cells (A), 4.5 μM IGFII antisense-treated cells (B), 4.5 μM IGFII sense-treated cells (C). X2 analysis showed the 4.5 μM IGFII antisense- induced apoptosis as mea- sured by the area under sub-G1 peak in the histograms were significant higher than that in the control and in the 4.5 μM IGFII sense- treated cells (P < 0.01). X-axis represents fluorescence intensity and Y-axis stands for relative cell numbers.
Discussion
One of direct method of detecting and quantifying apoptosis in cell cultures is to examine the morphology of cells after stained with fluorescent dyes such as PI6. A flow cytometry method based on a reduction in DNA staining due to cell shrinkage has been also widely used for detection of apoptosis10. Both methods have been used in this study to detect apoptotic effect of IGFII antisense in AO cells.
We have also attempted to detect the DNA fragmentation by gel electrophoresis (laddering) after IGFII antisense treatment of AO cells, but the results showed many smeared bands (data not shown). Although we could not offer the explanation for this, it has been reported that apoptosis occured in several systems without DNA laddering or with atypical DNA laddering11, and that in other systems necrosis cells also showed a distinct DNA ladder pattern12. In spite of this, our results, taken together as a whole, strongly suggest that IGFII antisense indeed prompts apoptosis in human cancer AO cells.
IGF-mediated myoblast survival was accompanied by stimulation of cell proliferation, as indicated by enhanced entry into S phase of the cell cycle and by increased cell number. IGFII, as an autocrine survival factor, was shown to be required for the survival of cultured hematopoietic cells after trophic factor withdrawal to prevent apoptosis, and to block death of a variety of tumor cell lines cultured for short term13. IGFII is also identified as the growth factor required for full tumorigenesis in transgenic mice expressing simian Virus 40 T antigen in the islets of Langerhans. In the absense of IGFII action, these islet cells show an enhanced rate of death and reduced tumor formation. In this study, it is further demonstrated that blocking IGFII expression using IGFII antisense oligonucleotides indeed promotes programmed cell death of human ovarian cancer AO cells.
S-oligonucleotides (phosphorothioate oligomers) have a sulfur residue instead of an oxygen residue linked to the phosphorus atom of the nucleotide backbone, they are considerably more resistant against eukaryotic endo- and exonucleases than unmodified single-stranded DNA, this is important for their use as prolonged gene suppressors. S-oligonucleotides have also been shown to be actively transported into the cell by a transporter protein. Most of the S-oligonucleotides appears to be located in the cytoplasm after extracellular application14. In summary, our results demonstrate that programmed cell death can be induced by inhibition of IGFII expression in human ovarian cancer AO cells, therefore, IGFII antisense could be a useful tool, with potential clinical application, to induce apoptosis in human ovarian cancer cells.
References
Langdon SP, Ritchie A, Young K, et al. Contrasting effects of 17 β-estradiol on the growth of human ovarian carcinoma cells in vitro and vivo. Int J Cancer 1993; 55:459–64.
Stewart CE, Rotwein P . Growth, differentiation, and survival: multiple physiological functions for insulin-like growth factors. Physiol Rev 1996;76(4):1005–26.
Ho MN, Delgado CH, Owens GA, Steller MA . Insulin-like growth factor-II participates in the biphasic effect of a gonadotropin-releasing hormone agonist on ovarian cancer cell growth. Fertil Steril 1997;67(5):870–6.
Bu SZ, Yin DL, Ren XH, et al. Progesterone induces apoptosis and up-regulation of p53 expression in human ovarian carcinoma cell lines. Cancer 1997; 79:1944–50.
Yin DL, Ren XH, Zheng ZL, et al. Etorphine inhibits cell growth and induces apoptosis in SK-N-SH cells: involvement of pertussis toxin-sensitive G proteins. Neurosci Research 1997;29(2):121–7.
Coucouvanis EC, Martin GR, Nadeau JH . Genetic approaches for studying programmed cell death during development of the laboratory mouse. In: Schwartz LM, Osborne BA. eds. Cell death. Methods in Cell Biology 1995; 46:397–9.
Yin DL, Ren XH, Bu SZ, et al. Induction of apoptosis and changes of bcl-2 expression in macrophage Ana-1 cells by all-trans retinoic acid. Cell Research 1996; 6:137–44.
Dolbeare F, Gratzner H, Pallavicinl MG, Gray JW . Flow cytometric measurement of total DNA content and incorporated bromodeoxyuridine. Proc Natl Acad Sci USA 1983; 80:5573–7.
McGahon AJ, Martin SJ, Bissonnette RP, et al. The end of the cell line: methods for the study of apoptosis in vitro In: Schwartz LM, Osborne BA. eds. Cell death. Methods in Cell Biology 1995; 46:165–9.
Telford WG, King LE, Fraker PJ . Rapid quantitation of apoptosis in pure and heterogeneous cell population using flow cytometry. J Immunol Methods 1994; 172:1–16.
Oberhammer F, Burseh W, Tiefenbacher R, et al. Apoptosis is induced by transforming growth factor-beta1 within 5 h in regressing liver without significant fragmentation of the DNA. Hepa- tology 1993; 18:1238–46.
Fukuda K, Kojiro M, Chiu JF . Demonstration of extensive chromatin cleavage in transplanted morris hepatoma 7777 tissue: apoptosis or necrosis? Am J Pathol 1993; 142:935–46.
Stewart CE, Rotwein P . Insulin-like growth factor-II is an autocrine survival factors for differ- entiating myoblasts. J Biol Chem 1996; 271:11330–8.
Schlingensiepen KH, Brysch W . Phosphorothioate Oligomers. In: Erickson RP, Izant JG. eds. Gene regulation: Biology of antisense RNA and DNA. Raven Press: New York 1992:317–9.
Acknowledgements
This research was supported by research grants from National Natural Science Foundation of China (No: 39600157), Chinese Academy of Sciences, Shanghai Research Centre of Life Science, and German Max-Planck Society. The authors thank Dr. Lan MA, Li-Zhen JIANG, Yan-Ping WANG for their help.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Yin, D., Pu, L. & Pei, G. Antisense oligonucleotide to insulin-like growth factor II induces apoptosis in human ovarian cancer AO cell line. Cell Res 8, 159–165 (1998). https://doi.org/10.1038/cr.1998.16
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cr.1998.16
Keywords
This article is cited by
-
Transcriptome comparisons detect new genes associated with apoptosis of cattle and buffaloes preantral follicles
Journal of Genetic Engineering and Biotechnology (2021)
-
Apoptosis in Granulosa cells during follicular atresia: relationship with steroids and insulin-like growth factors
Cell Research (2004)
