
Abstract
Oncolytic adenoviruses are under investigation as a promising novel strategy for cancer immunotherapeutics. Unfortunately, there is no immunocompetent mouse cancer model to test oncolytic adenovirus because murine cancer cells are generally unable to produce infectious viral progeny from human adenoviruses. We find that the murine K-ras-induced lung adenocarcinoma cell line ADS-12 supports adenoviral infection and generates infectious viral progeny. ADS-12 cells express the coxsackie and adenovirus receptor and infected ADS-12 cells express the viral protein E1A. We find that our previously described oncolytic virus, adenovirus TAV-255 (AdTAV-255), kills ADS-12 cells in a dose- and time-dependent manner. We investigated ADS-12 cells as an in-vivo model system for replicating oncolytic adenoviruses. Subcutaneous injection of ADS-12 cells into immunocompetent 129 mice led to tumor formation in all injected mice. Intratumoral injection of AdTAV-255 in established tumors causes a significant reduction in tumor growth. This model system represents the first fully immunocompetent mouse model for cancer treatment with replicating oncolytic adenoviruses, and therefore will be useful to study the therapeutic effect of oncolytic adenoviruses in general and particularly immunostimulatory viruses designed to evoke an antitumor immune response.
Similar content being viewed by others

Introduction
Oncolytic viruses preferentially replicate in cancer cells while sparing normal cells1 and can have a fundamental impact on cancer therapy.2 Whether replication-deficient or replication-competent, oncolytic viruses may provide selective and potent anticancer activity that can be due to viral replication or in some cases the expression of therapeutic transgenes.3 Adenovirus type 5 is a robust and flexible platform for gene delivery and has been modified to develop many oncolytic viruses. Unlike chemotherapeutic agents and molecularly targeted agents, oncoloytic viruses can replicate and can induce a potent immune response that can enhance the therapeutic activity of the virus, but can limit the distribution of the virus to tumor cells. Although repeated administration until progression is the norm for currently available antineoplastic agents, different strategies must be considered to maximize the potential of oncolytic viruses. In particular, the immune response to the virus, which is generally considered a barrier to repetitive administration, may be harnessed to enhance antitumor efficacy.
In order to model oncolytic adenovirus treatment, researchers have generally used human tumor xenografts that support viral replication in immunocompromised mice. Because xenografts require an immunodeficient host, those model systems will not reflect a host's adaptive immune response against the virus and cannot model the effect of the virus on inducing an antitumor immune response. To test the potential therapeutic activity of oncolytic viruses, an animal model should be immunocompetent and support active viral infection, including both cell lysis and production of infectious viral progeny. Unfortunately, mice are poor model systems for therapy with replication competent human adenoviruses because murine tumor cells are often not infected by human adenovirus and are generally unable to produce infectious viral progeny.4, 5 Consequently, our ability to study the impact of viral replication within tumor cells on the immune system is limited by lack of a mouse model system, particularly for adenovirus where human adenoviruses can infect mouse cells but do not complete an infective cycle to release new infectious particles.4, 5, 6, 7 In the absence of an effective mouse model system, researchers have turned to the Syrian hamster model;8 however, this model system, while effective for studying replicating human adenovirus, is limited by the accessibility of reagents to study immunological parameters. Therefore, the availability of a mouse model that could more effectively parallel the oncolytic activity of human adenoviruses could accelerate our ability to understand the interaction between oncolytic viruses and the immune system. In this study, we find that the murine lung adenocarcinoma cell line, ADS-12, supports adenoviral infection, expresses E1A, generates infectious viral progeny and responds to treatment with the oncolytic human adenovirus, AdTAV-255 (adenovirus TAV-255). Thus this novel K-ras mutant lung cancer model in fully immunocompetent mice is useful for evaluation of the host immune response to oncolytic human adenoviruses.
Materials and methods
Unless otherwise noted, all chemicals were purchased from Sigma-Aldrich (St Louis, MO, USA); cell culture reagents were obtained from Gibco (Grand Island, NY, USA) or Thermo Scientific (Waltham, MA, USA).
Cancer cell lines and adenovirus
Cancer cell lines
Murine K-ras mutant lung adenocarcinoma cell line (LKR-13) and murine sarcoma cell line (F244) were kindly provided Dr Tyler Jacks (Massachusetts Institute of Technology), and Dr Jack Bui (University of California, San Diego, CA, USA), respectively. ADR-12 cells were derived from LKR-13 cells by clonal isolation and testing for sensitivity to adenoviral infection. The murine melanoma cell line (B16F10), murine colorectal carcinoma cell line (CT26), human lung adenocarcinoma epithelial cell line (A549) and human embryonic kidney cell line (HEK293) were obtained from the American Type Culture Collection (ATCC). LKR-13, ADS-12 and F244 cells were grown in RPMI 1640 complete medium and CT26 cells and A549 cells were grown in Dulbecco’s modified Eagle’s complete medium. All complete medium were supplemented with 10% fetal calf serum, L-glutamine (2 mmol l−1), penicillin (100 IU ml−1), and streptomycin (50 μg ml−1). All cell lines were maintained at 37 °C in a humidified atmosphere at 5% CO2.
Adenovirus TAV-255
Adenovirus TAV-255 (AdTAV-255) carries a 50 nucleotide deletion in the promoter of E1A, possesses tumor selective expression of both E1a and E1b along with potent tumor-selective oncolytic activity.9 AdTAV-255 was produced in HEK-293A cells. Cells were infected with multiplicity of infection (MOI) of 1 and after 3 days post infection, cells were collected and resuspended in complete medium and lysed by three freeze-thaw cycles. Lysate was purified and concentrated using Fast-Trap Virus purification and concentration kits (Millipore, Billerica, MA, USA). Aliquots were kept frozen at −80 °C.
Virus plaque assay
Plaque assays were performed as described by VIRAPUR (San Diego, CA, USA). HEK-293 cells were seeded into six-well plates at 7.5 × 105. After 16 h, the cells were infected with 10 μl of each serial dilution of AdTAV-255 to the appropriately labeled well and incubated for 4 h in the incubator, then media was removed from the wells and they were overlaid with 2 ml 1% of SeaPlaque agarose in media. Plaques were visible by days 5–7 after infection. Wells were stained for 3 h at 37 °C by adding 100 μl of (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) MTT solution (5 mg ml−1 in phosphate-buffered saline (PBS)) and plaques were counted.
Cell growth curve
ADS-12 cells were plated on 12-well and 6-well plates at a concentration of 1 × 104 and 2 × 104 cells per well, respectively. These cells were allowed to grow for 3 days in Dulbecco’s modified Eagle’s complete medium supplemented with 10% fetal bovine serum, 2 mmol l−1 L-glutamine, 100 U ml−1 penicillin and 100 μg ml−1 streptomycin under a 5% CO2 atmosphere at 37 °C. The number of cells was counted every 24 h with trypsinization and trypan blue staining. Cell number was determined by using a hemocytometer. Cell viability was determined by trypan blue exclusion. Growth curve was generated by plotting the number of cells versus time.
Cell viability assay
MTT assay
ADS-12 cells and CT26 cells were seeded into 96-well plates at 5 × 103 or 1 × 104 cells per well, respectively. The next day they were infected with AdTAV-255 at MOI of 3, 10, 20 and 30 in triplicate based on cell number. At 3 and 6 days post infection, 10 μl MTT (5 mg ml−1) (Molecular Probes, Grand Island, NY, USA) was added into each well for 3 h, then 50 μl dimethyl sulfoxide was added to each well and the plates were incubated for 5 min at 37 °C to dissolve the formazan crystals. Absorbance was measured at a wavelength of 560/600 nm.
Crystal-violet staining
Cells were grown in 24-well plates and infected with AdTAV-255 at MOI of 5, 10 and 20. After 4 days post infection, the media was removed and cells were washed with PBS. Cells were then stained with crystal-violet (0.1% crystal-violet, 10% formaldehyde and 20% ethanol) for 20 min at room temperature, followed by rinsing with PBS and left for drying at room temperature.
Clonogenic assay
ADS-12 cells were seeded into 12-well plates at 2.5 × 104. The next day the cells were infected with AdTAV-255 at MOI of 10 and 20. After 3 days post infection, both uninfected and infected cells were counted and plated in six-well plates at 300 cells per well in duplicate. Once colonies were of sufficient size of at least 50 cells, culture medium was removed from the plates and cells were rinsed with PBS. Fixation and staining of clones was done with a mixture of crystal-violet (0.1% crystal-violet, 10% formaldehyde and 20% ethanol) for 20 min. Plates were rinsed with PBS and left for drying at room temperature.10 Counting of clones using Image J (imagej.nih.gov/ij) was done on the following day.
Viral yield assay
ADS-12, F244 and A549 cells were plated into 12-well plates at 1 × 105 cells per well. The next day, the cells were infected with AdTAV-255 at MOI of 10 based on cell number. At 48, 72 and 96 h post infection the plates were placed in −80 °C. The cells were lysed with three freeze thaw cycles to release virus. Viral titer from each sample was quantified by plaque assay using HEK-293A cells.
In a separate experiment, ADS-12 cells seeded at 2 × 105 cells per well were infected with TAV-255 at 1 MOI. Wells were supplemented with an additional 500 μl of media every 3 days. At 1, 4, and 7 days after infection, media was removed from triplicate wells without disturbing the monolayer, centrifuged for 1 min at 16 000 g, and the supernatant transferred to a new tube and stored at −80 °C until it was titered on HEK-293A cells with plaque assays.
Preparation of cell lysate and immunoblot analysis
Whole-cell lysate was prepared using M-PER mammalian protein extraction reagent (Pierce, Rockford, IL, USA). Protein concentration was determined using the NanoDrop ND-1000 spectrophotometer (Thermo Scientific). 25 μg of protein samples were treated and assayed on 4–12% NuPAGE Bis-Tris gels and transfered onto a polyvinylidene difluoride membrane according to the manufacturer's instructions. The membranes were blocked for 1 h at room temperature with blocking buffer and incubated overnight at 4 °C with primary antibodies in blocking buffer. The following antibodies were used: c-myc, the coxsackie and adenovirus receptor (CAR), E1A and β-tubulin. Proteins were visualized by enhanced chemiluminescence substrate.
Animals
129S4/SvJaeJ mice (stock no. 009104) mice were obtained from The Jackson Laboratory (Bar Harbor, ME, USA). Mice were bred and maintained at Moores Cancer Center Vivarium of University of San Diego according to guidelines established by the Institutional Animal Committee. 6- to 8-week-old male and female mice were used for all in vivo experiments.
Antitumor effects of oncolytic AdTAV-255
ADS-12 cells (5 × 105) in 100 μl PBS were injected subcutaneously in the right flanks of 129S4/SvJaeJ mice. When tumors reached mean value of 150 mm3, animals were randomized into two groups (PBS and AdTAV-255) and treatment was initiated. AdTAV-255 or PBS was given intratumorally (5 × 107 PFU per tumor in 50 μl PBS) every three days for three doses (on days 0, 3, and 6). Tumors were measured with a caliper every other day, and the tumor volume was calculated using the formula volume=(length × width2)/2. Mice were euthanized if their tumor diameter exceeded 1.5 cm or tumor volume exceeded 600 mm3. All animal procedures were approved by University of California, San Diego IACUC.
Statistical analysis
All determinations were performed in duplicate or triplicate; each experiment was repeated at least three times. Values are expressed as mean±s.e.m. Prism 4 software (GraphPad Software, La Jolla, CA, USA) was used for data analysis. Comparison between two groups was based on unpaired t-test. Log-rank test was performed to establish the significance of the differences between survival data.11 A value of P<0.05 was considered to be statistically significant.
Results
AdTAV-255 can infect and produce viral progeny in ADS-12 cells
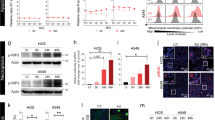
Human adenovirus type 5 infects and replicates poorly in murine cell lines.4, 5 We assessed if oncolytic AdTAV-255 can infect and replicate in murine ADS-12 cells. The CAR facilitates efficient viral uptake and is not present on all cell types.12, 13 E1A is the first viral gene product that is expressed during infection and is essential for viral replication and production.14 Thus, we first tested whether ADS-12 cells expresses CAR and if E1A is expressed after AdTAV-255 infection. We infected both ADS-12 cells and CT26 (mouse colon adenocarcinoma) cells with AdTAV-255 for 24, 48 and 72 h. Western blot analysis confirmed that ADS-12 cells express abundant CAR and CT26 cells lack CAR expression15 (Figure 1a). ADS-12 cells infected with the adenovirus AdTAV255 express the E1A protein within 24 h of infection, whereas CT26 cells have minimal expression at any time point (Figure 1a). We screened other murine cell lines and found heterogeneity in their permissiveness for E1A expression. F244 cells (murine sarcoma, which like ADS-12 cells are from a 129/Sv background) express E1A upon infection, while the murine melanoma cell line B16F10 which does not express CAR16 showed no E1A expression at the doses tested (Figure 1b). Many murine cells support E1A expression but are unable to support viral replication,17 so we next tested these cells for the ability to produce infectious progeny virions. We infected the mouse cell lines ADS-12, F244 and B16F10 for 72 and 96 h with 10 MOI of AdTAV-255 and performed viral titers after infection. Neither CT26 cells nor F244 cells produced viral progeny (data not shown). However, AdTAV-255 can replicate in ADS-12 cells in a time dependent manner (Figure 1c). Those experiments involved the usual practice of performing freeze-thaw cycles to release the virus, so we also tested whether the virus itself is able to induce lysis and escape from infected ADS-12 cells. We infected ADS-12 cells with AdTAV-255 at 1 MOI, supplemented the wells with media during the course of incubation to minimize cell death from starvation, collected media and centrifuged to remove cells and debris before titering. The number of plaque forming units in the media increased over time (Figure 1d), indicating that infectious viral progeny are released from ADS-12 cells after infection at low MOI without requiring further disruption. To ensure that the ADS-12 cell line we were using had not become contaminated with and overgrown by human cells, we isolated genomic DNA from the cells and amplified the lactate dehydrogenase gene. Sequencing this PCR product showed that it matched the mouse and not human isoform of the gene (data not shown). Thus, the ADS-12 cell line is a unique murine cell line that has CAR expression and supports the full lytic life cycle of adenovirus following AdTAV-255 infection.

Adenovirus TAV-255 (AdTAV-255) infects and replicates in ADS-12 cells. Western blot analysis of adenoviral E1A and CAR expression in different murine cell lines. (a) CT26 cells and ADS-12 cells were infected with AdTAV-255 at multiplicity of infection (MOI) of 10 for indicated times and assessed for E1A and CAR expression, with β-tubulin as a loading control. (b) Murine cell lines were infected with AdTAV-255 at MOI of 3 for 4 days and assessed for E1A expression. (c) ADS-12 cells were infected with AdTAV-255 at MOI of 10 for 72 and 96 h. The cells were collected and lysed, and total viral yields were determined with plaque assays. Each value is the average from three independent measurements. (d) ADS-12 cells were infected with AdTAV-255 at MOI of 1. Media was collected at 1, 4 and 7 days after infection without disturbing the cell monolayer, and the viral titer in the media was determined with plaque assays.
Our previous results demonstrated that AdTAV-255 had minimal cytotoxicity in nontransformed cells, but demonstrated potent cytolytic activity in a panel of human tumor cells including A549 cells.9 ADS-12 cells proliferate rapidly with a doubling time of 12–14 h compared with a doubling time of 24–35 h for other human and murine cancer cell lines (Figure 2a). We assessed the cytotoxicity of AdTAV-255 on a panel of murine tumor cell lines. First we compared ADS-12 cells with CAR negative CT26 cells. MTT assay results indicated that an MOI of 10 AdTAV-255 was highly lytic (80% cell death) in ADS-12 cells, whereas no cytotoxicity was observed in CT26 cells at 6 days post infection (Figure 2b). Crystal-violet staining showed that at an MOI of 10, AdTAV-255 induced ADS-12 cell death in 6 days. There was no cytotoxicity in other murine cancer cell lines such as B16F10 and F244 (Figure 2c). AdTAV-255 killed ADS-12 cells in a dose- and time-dependent manner on MTT assays (Figure 3a), and clonogenic assays showed that only a few colonies formed after infection at 10 MOI and no colonies formed at 20 MOI (Figure 3b).

TAV-255 induced cell death of ADS-12 cells but not other murine cells. (a) Growth of ADS-12 cells in cell culture. (b) ADS-12 cells and CT26 cells were infected with AdTAV-255 at multiplicity of infection (MOI) of 10 for 6 days and cell viability was assessed with MTT assays. (c) Crystal violet staining assays of ADS-12 cells and other murine cell lines infected with adenovirus TAV-255 (AdTAV-255) for 6 days.

Adenovirus TAV-255 (AdTAV-255) kills murine ADS-12 cells in a dose- and time-dependent manner. (a) ADS-12 cells were treated with different doses of AdTAV-255 for 3 and 6 days. Cell viability (expressed as a percentage related to the controls) was determined with MTT assays. (b) Clonogenic assays of ADS-12 cells infected with AdTAV-255. The graph shows relative numbers of colonies formed by ADS-12 cells after infection.
AdTAV-255 downregulates c-Myc expression in ADS-12 cells
c-Myc is frequently overexpressed in human cancers. The c-Myc oncoprotein is a transcription factor, with many of its target genes encoding proteins that control cell growth and proliferation.18, 19 In separate experiments, we found that adenoviral infection affects c-Myc expression in cancer cells, raising the possibility that adenovirus can be used to affect oncogenes that are not typically considered as ‘druggable’ targets (manuscript submitted). We tested A549 and ADS-12 cells for c-Myc expression at baseline and during infection with AdTAV-255. Our results indicated that AdTAV-255 downregulates c-Myc expression in both A549 and ADS-12 cell lines in time-dependent manner, and expression diminished within 72 h of infection (Figure 4).

Adenovirus TAV-255 (AdTAV-255) downregulates c-Myc expression in both ADS-12 and A549 cells. ADS-12 and A549 cells were infected with AdTAV-255 at an multiplicity of infection of 10. Lysates were collected at 24, 48 and 72 h post infection and probed for E1A and c-Myc.
AdTAV-255 inhibits subcutaneous tumor growth in immunocompetent mice
We next evaluated the oncolytic effect of AdTAV-255 on ADS-12 in vivo to assess its suitability as a preclinical model for therapeutic adenovirus. 5 × 105 ADS-12 cells were injected subcutaneously into 129S4/SvJaeJ mice. The injected cells formed tumors within 1 week. We then treated the mice with intratumoral injections of either AdTAV-255 at 5 × 107 PFU per dose or PBS, given every 3 days for three doses. AdTAV-255 inhibited tumor growth and led to significantly smaller tumor size in treated versus control mice 12 days after treatment (Figures 5a and b). Tumors in mice treated with three injections of TAV-255 were 278±45 mm3 (n=9) whereas tumors in mice treated with PBS were 531±66 mm3 (n=6; P<0.01). Viral treatment did not cause any discernible toxicity; both groups of mice were active until their tumors reached 1.5 cm in longest dimension or 600 mm3 volume, at which point they were euthanized. AdTAV-255 significantly improved survival in this cancer model (Figure 5c). 12 days after starting treatment, four out of six mice in the control group were sacrificed because of tumor size, but all mice in the AdTAV-255 group were alive.

Adenovirus TAV-255 (AdTAV-255) inhibits subcutaneous tumor growth in immunocompetent mice. 129S4/SvJaeJ mice were injected with 5 × 105 ADS-12 cells subcutaneously and allowed to form tumors. When the average tumor size reached ~150 mm3, ADS-12 tumors were treated with three intratumoral (i.t.) doses of AdTAV-255 (5 × 107 PFU per dose) or phosphate-buffered saline (PBS) every 3 days. (a) Mean tumor volume of each group is shown through day 12 after starting treatment, when four out of six mice treated with PBS were sacrificed due to tumor size. (b) Representative tumor images from PBS-treated and AdTAV-255-treated group. (c) A survival curve of mice. Mice were euthanized if their tumor diameter exceeded 1.5 cm or tumor volume exceeded 600 mm3. The quantitative data shown are the mean±s.e.m. *P<0.05, **P<0.01.
We did not observe any evidence of toxicity from the viral treatments, indicating that viral concentrations up to 5 × 107 PFU were well tolerated in these mice. These data suggest that significant amounts of AdTAV-255 could be administered without acute toxicity and that AdTAV-255 exhibited strong inhibitory effect on the growth of ADS-12 tumors.
Discussion
In this study, we find that the ADS-12 mouse lung cancer cell line is well suited for preclinical evaluation of oncolytic adenoviral therapy. ADS-12 cells express CAR, are killed by adenovirus and produce infectious viral progeny. ADS-12 cells are highly tumorigenic; tumors grew in all 15 of the injected mice and were sensitive to treatment with a human adenovirus. This allows us to, for the first time, model oncolytic adenoviral therapy in a fully immunocompetent mouse model.
A core feature of oncolytic viruses is the capacity to infect and kill tumor cells and release infectious viral progeny to subsequently infect and kill additional tumor cells.20 The potential for successive waves of tumor cell killing is dependent on replication of the virus and release of infectious viral progeny. A significant limitation of mouse model systems for studying oncolytic human adenoviruses is that, while many murine cell lines are infected by human adenoviruses and express viral proteins, they are unable to produce progeny virions.21 Therefore, to model the therapeutic impact of oncolytic viruses, it is critical to distinguish murine systems capable only of viral protein and DNA synthesis from those systems fully capable of producing infectious progeny virions. For example, a report by Wang et al.7 describes viral genome replication in mouse bladder transitional tumor cells and lung adenocarcinoma cells (LA795), but without production of infectious progeny virions. The term ‘viral replication assay’ has been applied to staining infected cells for expression of the viral protein hexon, which does occur in murine cells22 but does not establish that they produce infectious viral progeny as a plaque assay or limiting dilution assay does. One report described human adenovirus replication in a set of mouse cancer cell lines using limiting dilution assays,23 however, those findings could not be reproduced.24 Xenograft models have been used to study oncolytic adenoviruses in nude mice, but they cannot fully model the host immune response against both the virus and the cancer. Other model organisms have therefore been used including the Syrian hamster,25 cotton rats26, 27 and pigs.28 However, they lack the vast body of research and tools for studying immunologic responses available in mice.
ADS-12 cells were derived by clonal isolation and adenoviral sensitivity testing of LKR-13 lung adenocarcinoma cells isolated from a K-rasLA1 mouse.29, 30, 31 LKR-13 cells were used to study antitumoral role of curcumin and histone deacetylase inhibitor,32, 33 miRNA on tumorigenesis,34 and interaction between tumor cells and stromal cells.35 In this study, we demonstrate that ADS-12 cells express abundant levels of E1A protein, similar to the highly permissive human A549 cell line, and that the capacity of AdTAV-255 to suppress cellular oncogenes is preserved in the ADS-12 model. Therefore, the ADS-12 model will allow us to study the impact of gene expression changes in the tumor in the context of an immunologically competent model system.
Oncolytic viruses are showing efficacy in human clinical trials of JX-594 and talimogene iaherparepvec36 that use vaccinia and herpes viruses, respectively, to deliver granulocyte macrophage colony-stimulating factor. Further investigations into immunostimulatory viral therapies could be advanced by human adenoviruses in an immunocompetent mouse model system. For example, we have inserted tumor necrosis factor into the vector and can evaluate this activity of this vector in immunologically intact mice. We therefore anticipate that the ADS-12 model will be useful for preclinical studies of oncolytic adenoviruses, particularly immunomodulatory viruses, in a fully immunocompetent mouse. In summary, these data indicate that the ADS-12 cell line is a unique murine cancer cell line that supports infection by human adenovirus, expresses adenoviral proteins, undergoes lytic cell death and produces viable infectious particles following AdTAV-255 infection. This could therefore be used as a model system to study the effect of the host immune system on the vector-infected tumor, to evaluate the effects of oncolytic adenoviral therapy with an intact immune system, to determine the safety and biodistribution of replication-competent adenoviruses, modeling both the immune system's effects on inhibiting viral spread throughout the tumor and the potential for immunostimulatory viruses to induce an anticancer immune response.
References
Chiocca EA . Oncolytic viruses. Nat Rev Cancer 2002; 2: 938–950.
Heo J, Reid T, Ruo L, Breitbach CJ, Rose S, Bloomston M et al. Randomized dose-finding clinical trial of oncolytic immunotherapeutic vaccinia JX-594 in liver cancer. Nat Med 2013; 19: 329–336.
Vaha-Koskela MJ, Heikkila JE, Hinkkanen AE . Oncolytic viruses in cancer therapy. Cancer Lett 2007; 254: 178–216.
Duncan SJ, Gordon FC, Gregory DW, McPhie JL, Postlethwaite R, White R et al. Infection of mouse liver by human adenovirus type 5. J Gen Virol 1978; 40: 45–61.
Ginsberg HS, Moldawer LL, Sehgal PB, Redington M, Kilian PL, Chanock RM et al. A mouse model for investigating the molecular pathogenesis of adenovirus pneumonia. Proc Natl Acad Sci USA 1991; 88: 1651–1655.
Young AM, Archibald KM, Tookman LA, Pool A, Dudek K, Jones C et al. Failure of translation of human adenovirus mRNA in murine cancer cells can be partially overcome by L4-100K expression in vitro and in vivo. Mol Ther 2012; 20: 1676–1688.
Wang H, Wei F, Zhang J, Wang F, Li H, Chen X et al. A novel immunocompetent murine tumor model for the evaluation of RCAd-enhanced RDAd transduction efficacy. Tumour Biol 2012; 33: 1245–1253.
Thomas MA, Spencer JF, Wold WS . Use of the Syrian hamster as an animal model for oncolytic adenovirus vectors. Methods Mol Med 2007; 130: 169–183.
Hedjran F, Shantanu K, Tony R . Deletion analysis of Ad5 E1a transcriptional control region: impact on tumor-selective expression of E1a and E1b. Cancer Gene Ther 2011; 18: 717–723.
Franken NA, Rodermond HM, Stap J, Haveman J, van Bree C . Clonogenic assay of cells in vitro. Nat Protoc 2006; 1: 2315–2319.
Bortolanza S, Alzuguren P, Bunuales M, Qian C, Prieto J, Hernandez-Alcoceba R . Human adenovirus replicates in immunocompetent models of pancreatic cancer in Syrian hamsters. Hum Gene Ther 2007; 18: 681–690.
Bergelson JM, Cunningham JA, Droguett G, Kurt-Jones EA, Krithivas A, Hong JS et al. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science 1997; 275: 1320–1323.
Cohen CJ, Shieh JT, Pickles RJ, Okegawa T, Hsieh JT, Bergelson JM . The coxsackievirus and adenovirus receptor is a transmembrane component of the tight junction. Proc Natl Acad Sci USA 2001; 98: 15191–15196.
Deissler H, Opalka B . Therapeutic transfer of DNA encoding adenoviral E1A. Recent Pat Anticancer Drug Discov 2007; 2: 1–10.
Han SY, Lee YJ, Jung HI, Lee SW, Lim SJ, Hong SH et al. Gene transfer using liposome-complexed adenovirus seems to overcome limitations due to coxsackievirus and adenovirus receptor-deficiency of cancer cells, both in vitro and in vivo. Exp Mol Med 2008; 40: 427–434.
Mizuguchi H, Hayakawa T . Enhanced antitumor effect and reduced vector dissemination with fiber-modified adenovirus vectors expressing herpes simplex virus thymidine kinase. Cancer Gene Ther 2002; 9: 236–242.
Ganly I, Mautner V, Balmain A . Productive replication of human adenoviruses in mouse epidermal cells. J Virol 2000; 74: 2895–2899.
Prochownik EV . c-Myc as a therapeutic target in cancer. Expert Rev Anticancer Ther 2004; 4: 289–302.
Adhikary S, Eilers M . Transcriptional regulation and transformation by Myc proteins. Nat Rev Mol Cell Biol 2005; 6: 635–645.
Kirn D . Oncolytic virotherapy as a novel treatment platform for cancer. Ernst Schering Res Found Workshop 2003; 43: 89–105.
Lucher LA . Abortive adenovirus infection and host range determinants. Curr Top Microbiol Immunol 1995; 199: 119–152.
Zhang Z, Hu Z, Gupta J, Krimmel JD, Gerseny HM, Berg AF et al. Intravenous administration of adenoviruses targeting transforming growth factor beta signaling inhibits established bone metastases in 4T1 mouse mammary tumor model in an immunocompetent syngeneic host. Cancer Gene Ther 2012; 19: 630–636.
Hallden G, Hill R, Wang Y, Anand A, Liu TC, Lemoine NR et al. Novel immunocompetent murine tumor models for the assessment of replication-competent oncolytic adenovirus efficacy. Mol Ther 2003; 8: 412–424.
Ying B, Toth K, Spencer JF, Meyer J, Tollefson AE, Patra D et al. INGN 007, an oncolytic adenovirus vector, replicates in Syrian hamsters but not mice: comparison of biodistribution studies. Cancer Gene Ther 2009; 16: 625–637.
Thomas MA, Spencer JF, La Regina MC, Dhar D, Tollefson AE, Toth K et al. Syrian hamster as a permissive immunocompetent animal model for the study of oncolytic adenovirus vectors. Cancer Res 2006; 66: 1270–1276.
Toth K, Spencer JF, Tollefson AE, Kuppuswamy M, Doronin K, Lichtenstein DL et al. Cotton rat tumor model for the evaluation of oncolytic adenoviruses. Hum Gene Ther 2005; 16: 139–146.
Steel JC, Morrison BJ, Mannan P, Abu-Asab MS, Wildner O, Miles BK et al. Immunocompetent syngeneic cotton rat tumor models for the assessment of replication-competent oncolytic adenovirus. Virology 2007; 369: 131–142.
Jogler C, Hoffmann D, Theegarten D, Grunwald T, Uberla K, Wildner O . Replication properties of human adenovirus in vivo and in cultures of primary cells from different animal species. J Virol 2006; 80: 3549–3558.
Johnson L, Mercer K, Greenbaum D, Bronson RT, Crowley D, Tuveson DA et al. Somatic activation of the K-ras oncogene causes early onset lung cancer in mice. Nature 2001; 410: 1111–1116.
Wislez M, Fujimoto N, Izzo JG, Hanna AE, Cody DD, Langley RR et al. High expression of ligands for chemokine receptor CXCR2 in alveolar epithelial neoplasia induced by oncogenic kras. Cancer Res 2006; 66: 4198–4207.
Wislez M, Spencer ML, Izzo JG, Juroske DM, Balhara K, Cody DD et al. Inhibition of mammalian target of rapamycin reverses alveolar epithelial neoplasia induced by oncogenic K-ras. Cancer Res 2005; 65: 3226–3235.
Ma T, Galimberti F, Erkmen CP, Memoli V, Chinyengetere F, Sempere L et al. Comparing histone deacetylase inhibitor responses in genetically engineered mouse lung cancer models and a window of opportunity trial in patients with lung cancer. Mol Cancer Ther 2013; 12: 1545–1555.
Moghaddam SJ, Barta P, Mirabolfathinejad SG, Ammar-Aouchiche Z, Garza NT, Vo TT et al. Curcumin inhibits COPD-like airway inflammation and lung cancer progression in mice. Carcinogenesis 2009; 30: 1949–1956.
Kumar MS, Lu J, Mercer KL, Golub TR, Jacks T . Impaired microRNA processing enhances cellular transformation and tumorigenesis. Nat Genet 2007; 39: 673–677.
Zhong L, Roybal J, Chaerkady R, Zhang W, Choi K, Alvarez CA et al. Identification of secreted proteins that mediate cell-cell interactions in an in vitro model of the lung cancer microenvironment. Cancer Res 2008; 68: 7237–7245.
Senzer NN, Kaufman HL, Amatruda T, Nemunaitis M, Reid T, Daniels G et al. Phase II clinical trial of a granulocyte-macrophage colony-stimulating factor-encoding, second-generation oncolytic herpesvirus in patients with unresectable metastatic melanoma. J Clin Oncol 2009; 27: 5763–5771.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Zhang, L., Hedjran, F., Larson, C. et al. A novel immunocompetent murine model for replicating oncolytic adenoviral therapy. Cancer Gene Ther 22, 17–22 (2015). https://doi.org/10.1038/cgt.2014.64
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/cgt.2014.64
This article is cited by
-
Long-term antigen-specific immune response by an oncolytic adenovirus encoding SP-SA-E7-4-1BBL in HPV-16 cancer model
Molecular Biology Reports (2024)
-
Understanding and addressing barriers to successful adenovirus-based virotherapy for ovarian cancer
Cancer Gene Therapy (2021)
-
A new insight into aggregation of oncolytic adenovirus Ad5-delta-24-RGD during CsCl gradient ultracentrifugation
Scientific Reports (2021)
-
Defining a murine ovarian cancer model for the evaluation of conditionally-replicative adenovirus (CRAd) virotherapy agents
Journal of Ovarian Research (2019)
-
Whole cell vaccination using immunogenic cell death by an oncolytic adenovirus is effective against a colorectal cancer model
Molecular Therapy - Oncolytics (2016)
