
Abstract
Multiple myeloma (MM) is a malignant neoplasm of plasma cells. Although new molecular targeting agents against MM have been developed based on the better understanding of the underlying pathogenesis, MM still remains an incurable disease. We previously demonstrated that β-catenin, a downstream effector in the Wnt pathway, is a potential target in MM using RNA interference in an in vivo experimental mouse model. In this study, we have screened a library of more than 100 000 small-molecule chemical compounds for novel Wnt/β-catenin signaling inhibitors using a high-throughput transcriptional screening technology. We identified AV-65, which diminished β-catenin protein levels and T-cell factor transcriptional activity. AV-65 then decreased c-myc, cyclin D1 and survivin expression, resulting in the inhibition of MM cell proliferation through the apoptotic pathway. AV-65 treatment prolonged the survival of MM-bearing mice. These findings indicate that this compound represents a novel and attractive therapeutic agent against MM. This study also illustrates the potential of high-throughput transcriptional screening to identify candidates for anticancer drug discovery.
Similar content being viewed by others
Introduction
Multiple myeloma (MM) is a malignant neoplasm of plasma cells characterized by frequent chromosomal translocations and gene mutations.1 High-dose chemotherapeutics in conjunction with hematopoietic stem cell transplantation has been one standard clinical course of the treatment of MM.2, 3, 4 As we gain a better understanding of the pathogenesis underlying MM, new molecular targeting agents can be developed.5, 6 Nevertheless, MM remains incurable at present.7 Thus, it is important to continue to investigate new therapeutic agents that focus on targeting the biology of MM cells.
β-Catenin is a downstream effector in the canonical Wnt signaling pathway; thus, it is involved in the regulation of cell fate, proliferation and the self-renewal of stem cells. In the absence of Wnt ligands, cytosolic β-catenin undergoes proteasomal degradation through sequential phosphorylation and ubiquitination.8, 9 In the process, β-catenin that is phosphorylated by casein kinase I, and subsequently glycogen synthase kinase-3β, forms a degradation complex comprised of glycogen synthase kinase-3β, casein kinase I, axin and adenomatous polyposis coli. Phosphorylated β-catenin is ubiquitinated by cellular β-transducin repeat-containing proteins (β-TrCP), and subsequently degraded by the 26S proteasome.10 Upon Wnt ligand binding to Frizzled receptors and the co-receptor low-density lipoprotein receptor-related protein 5 (LRP5) or LRP6, the dishevelled phosphoprotein inactivates glycogen synthase kinase-3β, and the β-catenin degradation complex dissociates, resulting in the stabilization of β-catenin. Stabilized β-catenin translocates to the nucleus, where it binds to lymphoid enhancer-binding factor (LEF) and T-cell factor (TCF) proteins to activate the transcription of target genes such as c-myc and cyclin D1, which are involved in cell proliferation and survival.11, 12 Activation of Wnt signaling is closely linked to the process of carcinogenesis.13 Human MM-derived cell lines and primary MM cells overexpress β-catenin, whereas in normal plasma cells, β-catenin expression is undetectable.14, 15, 16 The recent study demonstrated that genes of soluble Wnt signal antagonists were methylated in MM cells, resulting in repression of their transcripts, and consequently, the increased expression of β-catenin.17 Overexpression of β-catenin promotes transcriptional activity of target genes related to the proliferation of MM cells, whereas inhibition of Wnt/β-catenin signaling suppresses MM growth.15, 16 These results suggest that β-catenin might be an attractive therapeutic target for MM.
Progress in molecular biology has enabled significant advances in drug discovery strategies. For example, RNA interference is a technique that can be used to knockdown the expression of specific genes in treated cells. An altered transcriptional profile can then be used as a biomarker for a specific molecular targeting, and quantitative evaluation can be applied to high-throughput technology.18, 19 Using high-throughput transcriptional screening technology (Supplementary Figure 1), we identified effective inhibitors of the Wnt/β-catenin signaling from a library of more than 100 000 chemical compounds (Supplementary Figure 2). From this initial series, we optimized a novel Wnt/β-catenin signal inhibitor, AV-65, that significantly suppressed the growth of MM cells and prolonged survival in a mouse model of orthotopic MM. These results demonstrate that AV-65 is a novel and attractive therapeutic agent for the treatment of MM.
Materials and methods
Cell lines, reagents and animals
Human AMO-1, KMS-12-BM, RPMI8226, NCI-H929, U226 and OPM-2 myeloma cell lines, and the IM-9 Epstein–Barr virus-transformed cell line derived from an MM patient, were purchased from the Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (Braunschweig, Germany). RPMI8226/LR5 melphalan-resistant cells were kindly provided by Dr William S Dalton (University of South Florida, Tampa, FL, USA).20 HCT-15 colorectal adenocarcinoma cells were purchased from American Tissue Culture Collection (Manassas, VA, USA). The 293T cells were purchased from the Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH. IM-9, RPMI8226, NCI-H929 and U266 cells were cultured in RPMI1640 (Gibco, Tokyo, Japan) containing 10% heat-inactivated fetal calf serum (FCS; Invitrogen, Tokyo, Japan), 2 mM L-glutamine (Gibco) and 1% penicillin–streptomycin (Gibco). The AMO-1 and KMS-12-BM cell lines were cultured in RPMI1640 containing 20% FCS, 2 mM L-glutamine and 1% penicillin–streptomycin. The RPMI8226/LR5 cell line was cultured in media containing 10% FCS, 2 mM L-glutamine, 1% penicillin–streptomycin and 5 μM melphalan (Sigma-Aldrich, Tokyo, Japan). Hepatocytes were cultured using CS-C complete medium kit (Cell Systems, Kirkland, WA, USA). HCT-15 cells were cultured in RPMI1640 containing 5% FCS, 2 mM L-glutamine and 1% penicillin–streptomycin. The 293T cells were cultured in Dulbecco's modified Eagle's medium containing 10% heat-inactivated fetal calf serum, 2 mM L-glutamine and 1% penicillin–streptomycin. All cell lines were maintained at 37 °C in a fully humidified atmosphere of 5% CO2 in air.
Peripheral blood mononuclear cells from three healthy volunteers were isolated by using Ficoll–Hypaque (GE Healthcare Japan, Tokyo, Japan), and cultured with in Iscove's modified Dulbecco's medium (Gibco) containing 20% FCS, 2 mM L-glutamine and 1% penicillin–streptomycin. Bone marrow (BM) samples were obtained from MM patients who were hospitalized in the Hospital of Kyoto Prefectural University of Medicine. In accordance with the Declaration of Helsinki recommendations, all procedures were approved by the institutional review board at Kyoto Prefectural University of Medicine, and written informed consent was obtained from every participant. Ficoll–Hypaque density centrifugation was used to separate mononuclear cells, followed by magnetic cell sorting (Miltenyi, Gladbach, Germany) using an anti-CD138 antibody (Ab) (Miltenyi) to enrich BM-derived MM cells. Cells were analyzed by FACS Canto II using the Diva software (BD Bioscience, Tokyo, Japan). The purity of the enriched CD138-positive cell populations was ⩾95%. Isolated MM cells were cultured in RPMI1640 containing 10% FCS, 2 mM L-glutamine and 20 ng/ml human interleukin-6 (R&D Systems, Minneapolis, MN, USA).
Specific pathogen-free 6- to 8-week old female nonobese diabetic-severe combined immune deficiency (NOD/SCID) mice (Clea Japan, Tokyo, Japan) were used for the in vivo experiments. Approval for these studies was obtained from the Committee on Animal Research of the Kyoto University Faculty of Medicine.
Growth inhibitory effects on myeloma cells
Cell proliferation was evaluated by the modified MTT (3-(4,5-dimethythiazol-2-yl)-2,5-diphenyl tetrazolium bromide) assay using Cell-Counting Kit-8 (Dojindo Laboratory, Kumamoto, Japan), as described previously.21, 22 Cells were seeded in a flat-bottomed 96-well plate (BD Bioscience) at a density of 3 × 103 cells in 100 μl of medium per well, and then incubated with serial dilutions of AV-65 for 72 h. The mean of four samples at each concentration was evaluated. Half maximal inhibitory concentration values were obtained using the nonlinear regression program CalcuSyn (Biosoft, Cambridge, UK).
Western blot analysis
Following treatment with AV-65, more than 1 × 106 cells were collected by centrifugation, and then the cells were washed with ice-cold phosphate-buffered saline (−) twice. Ice-cold radioimmunoprecipitation assay buffer (50 mM Tris-HCl (pH 7.4), 0.25 M NaCl, 5 mM EDTA, 20 mM NaF, 1% NP-40) containing fresh phenylmethylsulfonyl (1 mM) and protease inhibitor (10 μg/ml) was added to the cells. The suspension was transferred into a centrifuge tube and placed on ice for 15 min (min) with occasional vortexing to ensure complete lysis of the cells. The cell suspension was cleared by centrifugation at 14 000 g for 30 min at 4 °C. Nuclear and cytoplasmic protein fractions were obtained using NE-PER Nuclear and Cytoplasmic Extraction Reagents Kit (Pierce Biotechnology, Rockford, IL, USA), according to the manufacturer’s instructions. The supernatants (total cell lysate, nuclear and cytoplasmic protein fractions) were either used immediately or stored at −80 °C. Protein concentrations were determined using the DC Protein Assay (Bio-Rad Laboratories, Osaka, Japan). Immunoblotting was performed as described previously.16, 21 Samples (20 μg of protein) were analyzed using the following primary Abs, as indicated: anti-β-catenin (BD Pharmingen, San Jose, CA, USA), -Bad (Stressgen, Victoria, BC, Canada), -Bid (a kind gift from Dr David CS Huang, The Walter and Eliza Hall Institute of Medical Research (WEHI), Parkville, VIC, Australia),23 -Bim (clone 3C5, produced by Dr LA O’Reilly (WEHI)), -Bcl-2 (Bcl-2-100; Upstate, Lake Placid, NY, USA), -Bcl-xL (Stressgen), -Puma (ProSci, Poway, CA, USA), -Noxa (Alexis Biochemicals, Lausen, Switzerland), -Mcl-1 (Santa Cruz Biotechnology, Santa Cruz, CA, USA), -c-myc (Santa Cruz Biotechnology), -cyclin D1 (BD Pharmingen), -Oct-1 (Santa Cruz Biotechnology,), survivin (Cell Signaling Technology, Danvers, MA, USA) and -actin (Sigma-Aldrich). Horseradish peroxidase-coupled immunoglobulin G (Amersham Biosciences, Tokyo, Japan) was used as a secondary Ab, and immunoreactive proteins were detected by enhanced chemiluminescence or ECL-plus kits (Amersham Biosciences).
Ubiquitination of -catenin
At 12 h after AV-65 treatment, whole-cell lysates were obtained as described above. Lysates were subjected to immunoprecipitation using an anti-β-catenin monoclonal Ab (BD Pharmingen) and Dynabeads Protein A (Invitrogen), according to the manufacturer’s instructions. Ubiquitination of β-catenin was detected with anti-mono- and anti-poly-ubiquitinyl conjugates (Enzo Life Sciences International Inc., Plymouth Meeting, PA, USA).
TCF/LEF dual luciferase reporter assay
The activity of TCF/LEF transcription in HCT-15 cells was evaluated with the Wnt Cignal Reporter Assay (SABioscience, Fredrick, MD, USA). HCT-15 colorectal cancer cell line expresses high levels of β-catenin24 and is easily transfectable with plasmids. For each sample, 3 × 104 HCT-15 cells were reverse-transfected with 100 ng of a TCF/LEF firefly luciferase reporter plasmid and a constitutively expressing CMV-driven Renilla luciferase reporter with SureFECT Transfection Reagent (SABioscience, Fredrick, MD, USA), according to the manufacturer's instructions. At 16 h post-transfection, media were changed to assay media (Opti-MEM containing 0.5% FBS and 1% non-essential amino acids) for 8 h, followed by AV-65 treatment for 14 h. Relative luciferase activity of cells was detected using the Dual-Luciferase Reporter Assay System (Promega, Madison, WI, USA) and a Wallac Victor 1420 Plate Reader (Perkin-Elmer, Waltham, MA, USA). The relative luciferase activities generated by each well were calculated by determining the ratio of the luciferase (firefly/Renilla) values to normalize the signal. Three independent transfections were carried out in duplicate for each of the concentrations tested.
Induction of apoptosis and cell cycle analysis
Cell cycle analysis using propidium iodide (PI) was performed as described previously.22 Apoptosis induced by AV-65 was determined using Annexin-V-FITC Apoptosis Detection Kit I (BD Bioscience), according to the manufacturer’s instructions. Cells were analyzed by FACS Canto II using the Diva software (BD Bioscience). Caspase-3, -8 and -9 activities in the presence of AV-65 were evaluated using a fluorometric protease assay kit (MBL, Aichi, Japan), according to the manufacturer’s instructions.
Lentiviral transduction
We produced β-TrCP knockdown IM-9 cells using a lentivirus vector expressing short hairpin RNA against β-TrCP. Plasmids expressing short hairpin RNA against β-TrCP and green fluorescent protein were provided by Dr Kenzo Tokunaga (International Medical Center of Japan, Tokyo, Japan). The lentiviral packaging plasmid psPAX2 was provided by Addgene (Addgene plasmid 12260, D Trono, Lausanne, Switzerland). The pHIT/G plasmid25 expressing the vesicular stomatitis virus-G protein was kindly provided by M Malim (University of Pennsylvania, Philadelphia, PA, USA). In all, 10 μg of β-TrCP short hairpin RNA, 10 μg of psPAX2 and 10 μg of pHIT/G were added to a mixture of BES (N,N-bis[2-hydroxy-ethyl]-2-aminoethanesulfonic acid)-buffered saline and calcium chloride. After incubation for 20 min, the solution was added dropwise to a 10 cm dish of 70% confluent 293T cells. The cells were incubated for 12 h at 37 °C in a humidified incubator with an atmosphere of 3% CO2. The medium was replaced with 10 μM forskolin-containing medium and incubated at 37 °C and 10% CO2. After 48 h, the supernatants were harvested. In total, 1.0 × 106 IM-9 cells were transduced with 3 ml of the supernatant and polybrene (Sigma-Aldrich). We isolated GFP-positive cells by using a limiting dilution method.
Real-time polymerase chain reaction
RNA was purified with the QIAamp RNA Blood Mini Kit (Qiagen, Tokyo, Japan) and subjected to reverse transcription. Human β-TrCP gene expression levels were measured by a real-time polymerase chain reaction. A real-time polymerase chain reaction mixture contained 4 μl of Taqman master mix (Roche Diagnostics GmbH, Mannheim, Germany), cDNA, pairs of primers and Taqman probe (Universal Probe Library; Roche Diagnostics GmbH). The cDNA was amplified with a Light Cycler 3.5 (Roche Diagnostics GmbH) using the following parameters: 95 °C for 10 min, followed by 45 cycles of 95 °C for 15 s and 60 °C for 60 s. Glyceraldehyde-3-phosphate was used as an internal to normalize the loading difference. The primers for β-TrCP were as follows: forward primer, CCAGATAAATAACCATACACTGACCTC and reverse primer, CAGATACGTTAAATACCGCAACTTT. The primers for glyceraldehyde-3-phosphate were as follows: forward primer, AGCCACATCGCTCAGACAC and reverse primer, GCCCAATACGACCAAATCC.
In vivo effects of AV-65 on myeloma cells
The in vivo inhibitory effects of AV-65 on MM cells were investigated using an orthotropic mouse model. After exposure to 2 Gy of irradiation, specific pathogen-free 7- to 8-week old female NOD/SCID mice (Japan Clea, Osaka, Japan) were inoculated intravenously with 5 × 106 IM-9 cells in 100 μl of phosphate-buffered saline through the tail vein. IM-9 cells were successfully engrafted into the BM of all mice and IM-9-bearing mice died of MM approximately 32 days after inoculation. For the in vivo analysis of AV-65, the compound was formulated as a solid dispersion of 20%, the active drug ingredient with 80% of polyvinylpyrrolidine as an excipient to improve solubility. The in vivo formulation was administered intravenously through the tail vein in four cycles of 4 days on/1 day off, starting on the day following inoculation of IM-9 cells.
To investigate whether AV-65 treatment induced myelosuppression, we performed the colony-forming assay. After four cycles of AV treatment, NOD/SCID mice were killed and BM cells were obtained. Mononuclear cells were obtained by density centrifugation, and a colony-forming assay was performed using MethoCult GF M3434 (Stem Cell Technologies Inc., Vancouver, BC, Canada), according to the manufacturer’s instructions. The complete blood counts of the mice treated with vehicle or AV-65 were measured using the MEK-6450 Celltac α cell counter (Nihon Kohden, Tokyo, Japan).
Statistical analysis
The in vivo effects of AV-65 treatment were analyzed using the log-rank test; the effects of AV-65 on colony-forming ability and complete blood counts were analyzed using the Student’s t-test. A P-value of 0.05 was considered statistically significant.
Results
Inhibitory effects of AV-65 on myeloma cells in vitro
AV-65 inhibited growth of seven MM cell lines and the melphalan-resistant RMPMI8226/LR5 cell line in a dose-dependent manner (Figure 1a), with IC50 values after 72 h exposures in the range of 11.7–82.1 nM. However, AV-65 had lesser effects on normal peripheral blood mononuclear cells compared with MM cell lines (Figure 1a). AV-65 also inhibited the growth of IM-9 cells in a time-dependent manner (Figure 1b). CD138-positive myeloma cells obtained from MM patients were growth-inhibited by AV-65 in a dose-dependent manner (Figure 2c). Compared with the cell lines, higher doses of AV-65 were required for killing primary MM cells, presumably because the primary cells are quiescent and less sensitive to cytotoxic stimuli ex vivo.

Inhibition of myeloma cell proliferation. (a) The dose-dependent effect of compounds on the proliferation of myeloma cell lines was evaluated by the modified MTT assay. Cells were incubated with serial dilutions of compounds for 72 h. Data represent the means±s.d. of three independent experiments, and four replicate experiments. (b) Time-dependent inhibition of proliferation of IM-9 cells. Cells were incubated with AV-65 for 6, 12, 24 and 48 h. Data represent the means±s.d. of three independent experiments, and four replicate experiments. (c) Effect of compounds on CD138-positive myeloma cells and CD138-positive normal plasma cells. CD138-positive myeloma cells and CD138-normal plasma cells were purified from BM mononuclear cells obtained from myeloma patients and healthy volunteers, respectively. Data represent the mean of four replicates for each concentration.

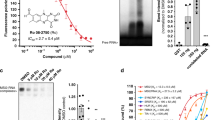
Alteration of β-catenin expression in myeloma cells by AV-65 treatment. (a) Cells were incubated with serial dilutions of AV-65 and harvested after 6, 12 and 18 h. Nuclear and cytoplasmic protein fractions were obtained and analyzed by western blot using an anti-β-catenin Ab. Results are representative of three independent experiments. (b) Ubiquitination of β-catenin by AV-65 treatment. Cells were incubated with the indicated concentration of AV-65 (1 × IC50), and then harvested after 12 h. Whole lysate was subjected to immunoprecipitation using an anti-β-catenin Ab, and immune complexes were captured using protein A beads. The protein A slurry (30 μl) was analyzed by immunoblot using anti-mono- and anti-poly-ubiquitinyl conjugates to detect ubiquitination of β-catenin (left panel). The bands of about 60 and 30 kDa detected in the membrane using anti-β-catenin Ab are nonspecific (right panel).
Alteration of -catenin expression and TCF transcriptional activity by AV-65
We next investigated alterations in β-catenin expression in MM cells. The levels of β-catenin in both the cytoplasmic and the nuclear fractions were diminished after 18 h by AV-65 treatment (Figure 2a). We also examined the ubiquitination of β-catenin in IM-9 cells following 12 h of exposure to AV-65. β-Catenin complexes were captured by immunoprecipitation with an anti-β-catenin Ab, and then analyzed by western blot using an Ab specific for ubiquitin–protein conjugates. Higher molecular weight ubiquitinated proteins were more prominent immunoprecipitates from AV-65-treated cells (Figure 2b), which suggested that AV-65 induces the degradation of β-catenin through the ubiquitin–proteasome pathway.
We next investigated the modulation of TCF transcriptional activity in AV-65-terated HCT-15 colorectal adenocarcinoma cells using a TCF/LEF dual luciferase reporter assay. The cells were transfected with a TCF/LEF firefly luciferase reporter plasmid along with a constitutively active CMV-driven Renilla luciferase reporter plasmid to ensure transfection efficiency. Transcriptional activity in HCT-15 cells was decreased by AV-65 treatment in a dose-dependent manner (Figure 3a). Moreover, the expression of cyclin D1, c-myc and survivin, which are downstream targets of TCF,11, 12, 26, 27, 28 were suppressed in IM-9 and U266 cells (Figure 3b). Taken together, these results suggest that AV-65 enhances the ubiquitination and subsequent proteasomal degradation of β-catenin, and that β-catenin/TCF target gene promoter activity is decreased.

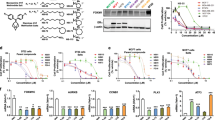
Inhibition of β-catenin-TCF signaling by AV-65 treatment. (a) Inhibition of TCF transcriptional activity in HCT-15 colorectal cancer cells. Cells were co-transfected with a TCF/LEF firefly luciferase reporter, and a CMV-driven Renilla luciferase reporter. To normalize transfection efficiency, cells were incubated with AV-65 for 14 h, and then luciferase activity was measured. Data represent the mean±s.d. of three independent experiments. The relative TCF transcription activity represents the difference between the relative luciferase units of treated versus non-treated cells. (b) Decreased expression of TCF downstream targets in IM-9 cells. Cells were incubated with serial dilutions of AV-65 and then harvested at 24 h. Harvested cells were lysed in radioimmunoprecipitation assay buffer. Cell extracts (20 μg of protein) was analyzed by western blot using the indicated Abs. Cyclin D1 and c-myc were not expressed in IM-9 and U266 cells, respectively.
-Catenin degradation via enhancement of -TrCP-mediated ubiquitination
We have clarified the mechanism by which AV-65 affects β-catenin degradation. β-Catenin is a target for the ubiquitin–proteasome pathway with β-TrCP, an ubiquitin ligase, triggering the ubiquitination of β-catenin. We examined the interaction between β-TrCP and β-catenin. The immunoprecipitation-western blotting analysis revealed that AV-65 enhanced the interaction of β-TrCP and β-catenin (Figure 4a).

Enhancement of the interaction between β-catenin and β-TrCP by AV-65 treatment. (a) Cells were incubated with serial dilutions of AV-65 and then harvested after 6 h. Whole lysate was subjected to immunoprecipitation using an anti-β-catenin Ab, and immune complexes were captured using protein A beads. The protein A slurry (30 μl) was analyzed by immunoblot using anti-β-TrCP Ab to detect binding between β-catenin and β-TrCP. (b) IM-9 cells were transduced with lentivirus-based short hairpin RNAs targeting β-TrCP and control vector. β-TrCP levels were measured by real-time polymerase chain reaction. Data represent the means±s.d. of three independent experiments, and four replicate experiments. (c) The alterations of β-catenin expression in IM-9 cells (left panel) and β-TrCP-KD IM-9 cells (right panel) by AV-65 treatment. After 24 h incubation with serial dilutions of AV-65, cells were harvested, and were lysed in radioimmunoprecipitation assay buffer. Cell extracts (20 μg of protein) were analyzed by western blot using an anti-β-catenin Ab and an anti-β-TrCP Ab.
We next established, using a lentivirus vector, a stable IM-9 cells containing a constitutive RNA interference capable of knockdown of β-TrCP (β-TrCP-KD IM-9) (Figure 4b). In contrast to IM-9 cells, the decrease of β-catenin was suppressed in β-TrCP-KD IM-9 cells after exposure to AV-65 (Figure 4c). In β-TrCP-KD IM-9 cells, AV-65-induced ubiquitination of β-catenin was suppressed (Supplementary Figure 3). These observations suggested that AV-65 degrades β-catenin through enhancing β-TrCP-mediated ubiquitination.
Induction of apoptosis by AV-65
Upon cell cycle analysis, we detected an increase in the sub-G1 phase fraction by AV-65 treatment in a dose- and a time-dependent manner (Supplementary Figures 4a and b, respectively). We also examined the induction of apoptosis in AV-65-treated cells by Annexin-V/propidium iodide staining. Early apoptotic cells (Annexin-V-positive/propidium iodide -negative), late apoptotic cells and necrotic cells (Annexin-V-positive/propidium iodide -positive) increased in a dose- and time-dependent manner (Figure 5a and Supplementary Figure 5, respectively). These results indicated that AV-65 inhibits the proliferation of MM cells through the induction of apoptosis.

Induction of apoptosis in myeloma cells by AV-65. (a) Cells were incubated with serial dilutions of AV-65 and harvested after 24 h. Data are representative of three independent experiments. The numbers inside each quadrant indicate the percentage of the cell population with the quadrant characteristic. (b) Cells were incubated with serial dilutions of AV-65 and harvested after 24 h. Harvested cells were lysed in lysis buffer, and then caspase-3, -8 and -9 activity was evaluated using a fluorometric protease assay. Data represent the means±s.d. of three independent experiments. Solid, gray and open bars indicate caspase-3, -8 and -9, respectively.
We next investigated the underlying mechanism of apoptosis induced by AV-65 using a fluorometric protease assay. AV-65 treatment activated caspase-3, caspase-8 and caspase-9, indicating that apoptosis by AV-65 involves the activation of both intrinsic and extrinsic apoptotic pathways (Figure 5b). Western blot analysis showed that the expression of Noxa was increased in IM-9, AMO-1 and OPM-2 cells by AV-65 treatment, and the expression of Bad and BimS was increased in U266 cells. In AMO-1 cells, tBid and Noxa levels were increased, and the expression of Noxa was increased in OPM-2 cells. No significant changes in the expression of antiapoptotic proteins such as Bcl-2, BCl-xL and the long isoform of Mcl-1 were observed (Supplementary Figure 6). These findings suggested that AV-65 induces apoptosis accompanied with the activation of caspases through the cell type-dependent activation of proapoptotic BH-3-only proteins.
Inhibitory effects on myeloma in vivo
We assessed the in vivo effects of AV-65 on the growth of MM cells in vivo in an orthotopic mouse model of MM. Following engraftment of IM-9 cells into NOD/SCID mice, all animals died of MM by approximately 32 days after transplantation (Figure 6a and Supplementary Figure 7). When we administered AV-65 to MM-bearing mice intravenously, the survival of mice treated with the higher doses of AV-65 was significantly prolonged as compared to vehicle-treated mice (P=0.028; Figure 6a). To investigate the adverse effects of AV-65 in treated mice, we assayed colony-forming activity in BM and other biochemical parameters in AV-65 and vehicle-treated mice. There were no statistical differences in body weight (data not shown), the complete blood cell counts between the groups (Figure 6b) or the numbers of colony-forming units in BM (Figure 6c). These observations indicated that AV-65 is a potent therapeutic agent for MM and does not induce severe adverse effects.

The in vivo inhibitory effects of AV-65 on MM cells in an orthotopic mouse model. Following irradiation (2 Gy), specific pathogen-free 7- to 8-week old female NOD/SCID mice were inoculated intravenously with 5 × 106 IM-9 cells in 100 μl phosphate-buffered saline through the tail vein. The day after inoculation, AV-65 was administered intravenously for four cycles of 4 days on/1 day off. (a) Survival of IM-9-bearing mice treated with higher doses of AV-65 was significantly prolonged compared with vehicle-treated mice (P=0.028). Red, green and blue lines represent the survival rates of high dose of AV-65 (8 mg/kg), low dose of AV-65 (4 mg/kg) and vehicle-treated groups, respectively (10 mice per group). (b) The complete blood counts of vehicle- of AV-65-treated mice. There was no significant difference between mice treated with the higher doses of AV-65 and vehicle. Data represent the means±s.d. of four mice in each group. (c) Influence on BM hematopoiesis in treated mice. After four courses of AV-65 treatment, NOD/SCID mice were killed, and BM cells were obtained. Mononuclear cells were obtained by density centrifugation and analyzed by colony-forming assay. Colony-forming assay was performed in duplicate for each mouse (4 mice per group). There was no significant difference between mice treated with the higher doses of AV-65 and vehicle. Data represent the means±s.e. of four mice in each group.
Discussion
Over the past 20 years, drug discovery has been dominated by phenotypic screening based on cancer cell biology. Using a traditional, pharmacology-driven approach, compounds that directly affected the pathophysiology of cancer cells were identified. Advances in molecular biology and genetic engineering has brought out the era of chemical genetics,29 leading to well-characterized selective cancer-specific drugs such as imatinib and trastuzumab, which target specific molecular pathways. In recent years, the Human Genome Project and related efforts have led to the establishment of databases based on genomics and transcriptome analyses, and transcriptional profiling and high-throughput technology using microarrays have been applied to systemic drug screening.30 Profiling the expression of numerous gene sets in normal versus pathological states can provide critical clues to intracellular signaling pathway function, as well as to identities of genes that play important roles in carcinogenesis.
β-Catenin is a downstream effector of the canonical Wnt signaling pathway. The activation of Wnt signaling is closely linked to carcinogenesis,13, 31, 32 and β-catenin is considered a valid drug target known to regulate the Wnt pathway.27, 33 In support of this, downregulation of β-catenin by small interfering RNA inhibits the proliferation of cancer cells.16, 34, 35 Recently, a study describing the high-throughput transcriptional screening analysis (Supplementary Figure 1) of HT29 colon adenocarcinoma cells transfected with an small interfering RNA against β-catenin was reported.18 In HT-29 cells, β-catenin overexpression correlates with carcinogenesis due to mutation of adenomatous polyposis coli,11 making these cells a useful model system. Transcriptional changes before and after knockdown of β-catenin were profiled using a gene expression microarray, and nine genes (CTNNB1, NM_001904; CEACAM6, NM_002483; EMP1, NM_001423; IHH, NM_002181; MYEF2, NM_016132; NTN4, NM_021229; DHRS9, NM_005771; HMGB1, NM_002128; and GAPDH, NM_002046; GAPDH was used as control) were identified as candidate genes that were modulated by inhibition of the Wnt/β-catenin pathway. The gene set was then validated using a series of small-molecule probes that are known inhibitors of the Wnt/β-catenin signaling (that is, indomethacin and retinoic acid).36, 37, 38 A random forest algorithm developed during the signature validation phase was used to identify hit compounds from the screening library. Hit compounds and closely related analogs were re-tested in escalating doses to confirm activity and measure potency, and several early-stage compound series was selected for further analysis as inhibitors of the Wnt/β-catenin pathway. Ultimately, the LC-363 compound series (Supplementary Figure 2) were selected for further analysis as an inhibitor of the Wnt/β-catenin pathway. AV-65 was generated based on initial LC-363 series. Our preliminary data show that this AV-65 compound was more potent in inhibiting the Wnt/β-catenin signaling than the early-stage compound series (data not shown). Lepourcelet et al.39 screened several antagonistic compounds for their ability to inhibit the β-catenin pathway using a high-throughput method for immunoenzymatic detection based on protein–protein interactions. The screened compounds were from natural sources, and several of them shared polyhydroxylated planar features. One of these compounds, PKF115-584, inhibited the proliferation of MM cells.15 SDX-308, an etodolac analog, also induced cytotoxicity in MM cells through inhibition of the β-catenin pathway.40 By comparison, this novel compound of this study is fundamentally different from these earlier compounds in both structure and function.
We first examined the in vitro effects of AV-65 on MM cells. AV-65 inhibited the proliferation of MM cells in a dose- and a time-dependent manner (Figures 1a and b, respectively). Moreover, β-catenin was degraded in AV-65-treated cells through an increase in ubiquitination (Figure 2b). The results of a dual luciferase reporter assay indicated that AV-65 inhibits TCF transcriptional activity, and that c-myc, cyclin D1 and survivin expression is decreased by AV-65 (Figures 3a and b). Moreover, β-catenin was degraded in AV-65-treated cells through an increase in ubiquitination (Figure 2b). Moreover, AV-65 enhanced the interaction between β-catenin and β-TrCP, resulting in the increased ubiquitination and degradation of β-catenin. The involvement of β-TrCP was verified by experiments showing knockdown of β-TrCP (Figure 4). AV-65 inhibited TCF/LEF transcriptional activity, resulting in a decrease in the level of TCF downstream targets and the induction of apoptosis through the activation of caspases (Figures 5a and b). Thus, AV-65 inhibits the Wnt/β-catenin signaling through the decrease of β-catenin/TCF transcriptional activity, resulting in the suppression of MM cell proliferation.
We also investigated the in vivo effects of a water-soluble formulation of AV-65. Survival of MM-bearing mice was significantly prolonged by administration of AV-65 (Figure 6a). AV-65 had no adverse effects on the mice, in terms of body weight (data not shown), the number of the peripheral blood cells or the colony-forming units in the BM (Figures 6b and c). We are planning to carry out a phase I study with this compound series in solid and hematopoietic malignancies.
In conclusion, a novel Wnt/β-catenin signaling inhibitor, AV-65, inhibited the proliferation of MM cells in vivo and in vitro. Thus, β-catenin represents a promising molecular target for therapy against MM, and AV-65 a promising agent against MM.
References
Kuehl WM, Bergsagel PL . Multiple myeloma: evolving genetic events and host interactions. Nat Rev Cancer 2002; 2: 175–187.
Harousseau JL, Moreau P . Autologous hematopoietic stem-cell transplantation for multiple myeloma. N Engl J Med 2009; 360: 2645–2654.
Moreau P, Hullin C, Garban F, Yakoub-Agha I, Benboubker L, Attal M et al. Tandem autologous stem cell transplantation in high-risk de novo multiple myeloma: final results of the prospective and randomized IFM 99-04 protocol. Blood 2006; 107: 397–403.
Rosinol L, Perez-Simon JA, Sureda A, de la Rubia J, de Arriba F, Lahuerta JJ et al. A prospective PETHEMA study of tandem autologous transplantation versus autograft followed by reduced-intensity conditioning allogeneic transplantation in newly diagnosed multiple myeloma. Blood 2008; 112: 3591–3593.
Bringhen S, Avonto I, Magarotto V, Boccadoro M, Palumbo A . Investigational treatments for multiple myeloma. Expert Opin Investig Drugs 2006; 15: 1565–1582.
Hideshima T, Mitsiades C, Tonon G, Richardson PG, Anderson KC . Understanding multiple myeloma pathogenesis in the bone marrow to identify new therapeutic targets. Nat Rev Cancer 2007; 7: 585–598.
Rajkumar SV . Treatment of myeloma: cure vs control. Mayo Clin Proc 2008; 83: 1142–1145.
Aberle H, Bauer A, Stappert J, Kispert A, Kemler R . Beta-catenin is a target for the ubiquitin–proteasome pathway. EMBO J 1997; 16: 3797–3804.
Kikuchi A . Regulation of beta-catenin signaling in the Wnt pathway. Biochem Biophys Res Commun 2000; 268: 243–248.
Hacein-Bey-Abina S, Von Kalle C, Schmidt M, McCormack MP, Wulffraat N, Leboulch P et al. LMO2-associated clonal T cell proliferation in two patients after gene therapy for SCID-X1. Science 2003; 302: 415–419.
He TC, Sparks AB, Rago C, Hermeking H, Zawel L, da Costa LT et al. Identification of c-MYC as a target of the APC pathway. Science 1998; 281: 1509–1512.
Tetsu O, McCormick F . Beta-catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature 1999; 398: 422–426.
Katoh M, Katoh M . WNT signaling pathway and stem cell signaling network. Clin Cancer Res 2007; 13: 4042–4045.
Derksen PW, Tjin E, Meijer HP, Klok MD, MacGillavry HD, van Oers MH et al. Illegitimate WNT signaling promotes proliferation of multiple myeloma cells. Proc Natl Acad Sci USA 2004; 101: 6122–6127.
Sukhdeo K, Mani M, Zhang Y, Dutta J, Yasui H, Rooney MD et al. Targeting the beta-catenin/TCF transcriptional complex in the treatment of multiple myeloma. Proc Natl Acad Sci USA 2007; 104: 7516–7521.
Ashihara E, Kawata E, Nakagawa Y, Shimazaski C, Kuroda J, Taniguchi K et al. {beta}-Catenin small interfering RNA successfully suppressed progression of multiple myeloma in a mouse model. Clin Cancer Res 2009; 15: 2731–2738.
Chim CS, Pang R, Fung TK, Choi CL, Liang R . Epigenetic dysregulation of Wnt signaling pathway in multiple myeloma. Leukemia 2007; 21: 2527–2536.
Bol D, Ebner R . Gene expression profiling in the discovery, optimization and development of novel drugs: one universal screening platform. Pharmacogenomics 2006; 7: 227–235.
Strand KJ, Khalak H, Strovel JW, Ebner R, Augustus M . Expression biomarkers for clinical efficacy and outcome prediction in cancer. Pharmacogenomics 2006; 7: 105–115.
Dalton WS, Durie BG, Alberts DS, Gerlach JH, Cress AE . Characterization of a new drug-resistant human myeloma cell line that expresses P-glycoprotein. Cancer Res 1986; 46: 5125–5130.
Kuroda J, Kimura S, Segawa H, Kobayashi Y, Yoshikawa T, Urasaki Y et al. The third-generation bisphosphonate zoledronate synergistically augments the anti-Ph+ leukemia activity of imatinib mesylate. Blood 2003; 102: 2229–2235.
Kawata E, Ashihara E, Kimura S, Takenaka K, Sato K, Tanaka R et al. Administration of PLK-1 small interfering RNA with atelocollagen prevents the growth of liver metastases of lung cancer. Mol Cancer Ther 2008; 7: 2904–2912.
Kaufmann T, Tai L, Ekert PG, Huang DC, Norris F, Lindemann RK et al. The BH3-only protein bid is dispensable for DNA damage- and replicative stress-induced apoptosis or cell-cycle arrest. Cell 2007; 129: 423–433.
Rice PL, Kelloff J, Sullivan H, Driggers LJ, Beard KS, Kuwada S et al. Sulindac metabolites induce caspase- and proteasome-dependent degradation of beta-catenin protein in human colon cancer cells. Mol Cancer Ther 2003; 2: 885–892.
Fouchier RA, Meyer BE, Simon JH, Fischer U, Malim MH . HIV-1 infection of non-dividing cells: evidence that the amino-terminal basic region of the viral matrix protein is important for Gag processing but not for post-entry nuclear import. EMBO J 1997; 16: 4531–4539.
van de Wetering M, Oving I, Muncan V, Pon Fong MT, Brantjes H, van Leenen D et al. Specific inhibition of gene expression using a stably integrated, inducible small-interfering-RNA vector. EMBO Rep 2003; 4: 609–615.
Dihlmann S, von Knebel Doeberitz M . Wnt/beta-catenin-pathway as a molecular target for future anti-cancer therapeutics. Int J Cancer 2005; 113: 515–524.
Zhang T, Otevrel T, Gao Z, Gao Z, Ehrlich SM, Fields JZ et al. Evidence that APC regulates survivin expression: a possible mechanism contributing to the stem cell origin of colon cancer. Cancer Res 2001; 61: 8664–8667.
Stockwell BR . Chemical genetics: ligand-based discovery of gene function. Nat Rev Genet 2000; 1: 116–125.
Spring DR . Chemical genetics to chemical genomics: small molecules offer big insights. Chem Soc Rev 2005; 34: 472–482.
Polakis P . The many ways of Wnt in cancer. Curr Opin Genet Dev 2007; 17: 45–51.
Moon RT, Kohn AD, De Ferrari GV, Kaykas A . WNT and beta-catenin signalling: diseases and therapies. Nat Rev Genet 2004; 5: 691–701.
Daniels DL, Eklof Spink K, Weis WI . Beta-catenin: molecular plasticity and drug design. Trends Biochem Sci 2001; 26: 672–678.
Verma UN, Surabhi RM, Schmaltieg A, Becerra C, Gaynor RB . Small interfering RNAs directed against beta-catenin inhibit the in vitro and in vivo growth of colon cancer cells. Clin Cancer Res 2003; 9: 1291–1300.
Huang WS, Wang JP, Wang T, Fang JY, Lan P, Ma JP . ShRNA-mediated gene silencing of beta-catenin inhibits growth of human colon cancer cells. World J Gastroenterol 2007; 13: 6581–6587.
Dihlmann S, Siermann A, von Knebel Doeberitz M . The nonsteroidal anti-inflammatory drugs aspirin and indomethacin attenuate beta-catenin/TCF-4 signaling. Oncogene 2001; 20: 645–653.
Hawcroft G, D’Amico M, Albanese C, Markham AF, Pestell RG, Hull MA . Indomethacin induces differential expression of beta-catenin, gamma-catenin and T-cell factor target genes in human colorectal cancer cells. Carcinogenesis 2002; 23: 107–114.
Easwaran V, Pishvaian M, Salimuddin, Byers S . Cross-regulation of beta-catenin-LEF/TCF and retinoid signaling pathways. Curr Biol 1999; 9: 1415–1418.
Lepourcelet M, Chen YN, France DS, Wang H, Crews P, Petersen F et al. Small-molecule antagonists of the oncogenic Tcf/beta-catenin protein complex. Cancer Cell 2004; 5: 91–102.
Yasui H, Hideshima T, Ikeda H, Ocio EM, Kiziltepe T, Vallet S et al. Novel etodolac analog SDX-308 (CEP-18082) induces cytotoxicity in multiple myeloma cells associated with inhibition of beta-catenin/TCF pathway. Leukemia 2007; 21: 535–540.
Acknowledgements
This work was partly supported by Grant-in-Aids for Scientific Research from the Ministry of the Education, Culture, Sports, Science and Technology of Japan (EA, SK, HH and TM).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
H Yao, E Ashihara, Y Nakagawa, J Kuroda, R Nagao, R Tanaka, A Yokota, M Takeuchi, K Sakai, C Shimazaki, M Taniwaki, H Hirai, S Kimura and Taira Maekawa disclose no financial conflict of interest. JW Strovel, Kathyrn Strand and Janak Padia are employees of PGx Health, a Division of Clinical Data Inc.
Additional information
Supplementary Information accompanies the paper on Blood Cancer Journal website
Supplementary information
Rights and permissions
This work is licensed under the Creative Commons Attribution-NonCommercial-No Derivative Works 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Yao, H., Ashihara, E., Strovel, J. et al. AV-65, a novel Wnt/β-catenin signal inhibitor, successfully suppresses progression of multiple myeloma in a mouse model. Blood Cancer Journal 1, e43 (2011). https://doi.org/10.1038/bcj.2011.41
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/bcj.2011.41
Keywords
This article is cited by
-
The Wnt/β-catenin signalling pathway in Haematological Neoplasms
Biomarker Research (2022)
-
Smenospongidine suppresses the proliferation of multiple myeloma cells by promoting CCAAT/enhancer-binding protein homologous protein-mediated β-catenin degradation
Archives of Pharmacal Research (2017)
-
Targeting of BMI-1 with PTC-209 shows potent anti-myeloma activity and impairs the tumour microenvironment
Journal of Hematology & Oncology (2016)
-
Signaling Pathways and Emerging Therapies in Multiple Myeloma
Current Hematologic Malignancy Reports (2016)
-
High expression of endoplasmic reticulum chaperone grp94 is a novel molecular hallmark of malignant plasma cells in multiple myeloma
Journal of Hematology & Oncology (2015)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}