
Abstract
Extracellular vesicles (EVs) comprise apoptotic bodies, microvesicles and exosomes, and they perform as key regulators in cell-to-cell communication in normal as well as diseased states. EVs contain natural cargo molecules, such as miRNA, mRNA and proteins, and transfer these functional cargos to neighboring cells or more distant cells through circulation. These functionally active molecules then affect distinct signaling cascades. The message conveyed to the recipient cells is dependent upon the composition of the EV, which is determined by the parent cell and the EV biogenesis. Because of their properties such as increased stability in circulation, biocompatibility, low immunogenicity and toxicity, EVs have drawn attention as attractive delivery systems for therapeutics. This review focuses on the functional use of exosomes in therapy and the potential advantages and challenges in using exosomes for therapeutic purposes.
Similar content being viewed by others

Introduction
Recently, extracellular vesicles (EVs) have earned a significant contribution in understanding and targeting different diseases. Over the last few years, many studies have been performed in the field of microvescicles to study their role in regulation of different pathophysiological conditions, such as cancer1, immunological disorders2, and other systematic disorders in different organs. Furthermore, recent research has been performed to understand the impact of stem cell-derived microvesicles in therapeutic intervention and tissue repair3. EVs are membrane-bound endocytic vesicles released in an evolutionally conserved manner by cells ranging from organisms such as prokaryotes to higher eukaryotes and plants that are capable of intercellular communication. Due to their capacity to transfer proteins, lipids and nucleic acids, they can influence various physiological and pathological functions of both recipient and parent cells4.
Recently, several studies have demonstrated that these membrane-bound vesicles serve as biomarkers to identify different diseases and are essential for effectively combating disease. Moreover, due to their ability to carry biological materials to tissues, exosomes have vast application potential in therapy. Nonetheless, many concerns have arisen regarding the therapeutic use of stem cells, such as undesirable proliferation, heterogeneity and immunogenic effects. In this context, stem cell-derived cell-free exosomes are considered a major emerging tool for curing many diseases. Previously, we discussed the major contents and paracrine-mediated functions of exosomes. These exosomes have both beneficial physiological and detrimental pathological roles during their biological processes. Moreover, studies have shown that exosomes are carriers for different micro-RNAs (miRNAs) that are responsible for different cardiac diseases, including hypertension, cardiac hypertrophy, cerebral ischemia (CI) and stroke5,6. This review gives a brief overview of studies that have been reported regarding how exosomes serve as a new molecular target for various diseases, and it discusses the use of exosomes in therapy.
Classification
EVs are small membrane vesicles heterogeneous in size (20 nm to 2 μm) bounded by a phospholipid bilayer and released by all cell types in various biological fluids and extracellular space7. They are released through exocytic budding of the plasma membrane in response to cellular activation or apoptosis8. These vesicles contain membrane surface protein and lipid components as well as nucleic acids originating from their original cells4. EVs can be classified into different subpopulations, including apoptotic bodies (ABs), microvesicles (MVs) and exosomes, each with specific characteristics, including their biogenesis, size, cellular origin, protein composition, mRNA and miRNA content, and/or biological function (Table 1)9.
Exosomes
The meaning of the term exosomes has changed over time in efforts to better understand their endocytic origin and distinguish them from other microvesicles. Exosomes are currently defined as heterogeneous molecules with a diameter of less than 100 nm and a density of 1.10–1.18 g/mL. They also reflect the phenotypic state of the parent cells from which they are generated10. Exosomes originate from repeated invagination of the lipid bilayer membrane of multivesicular bodies, and at any given point, they may contain all known molecular constituents of a cell, including proteins, RNA, and DNA11. The fusion of multivesicular bodies with the plasma membrane facilitates the release of exosomes into the extracellular space.
Microvesicles
Microvesicles, described by Wolf for the first time in 1967, have been studied for the past two decades12. Microparticles or microvesicles (MVs) originate directly from the membrane surface through outward budding of the plasma membrane. They are characterized by the presence of phosphatidylserine (PS) in the outer leaflet of the membrane and a size ranging between 100 and 1000 nm in diameter9. MVs are heterogeneous populations with irregular shape and varied size and densities between 1.04 and 1.07 g/mL9,13. MVs can be separated from different biological samples by size along with a combination of typical cell-specific markers, which will also help to detect their cellular origin9. At present, less information has been found for markers to identify MVs compared with exosomes. However, the most common MV markers used thus far in studies are CD40 ligand, adenosine diphosphate ribosylation factor 6, and several integrins and selectins14. MV inner contents are similar to those of exosomes and are mirror those of their parent cells.
Apoptotic bodies
In 1974, Kerr first described apoptotic bodies as vesicle-like structures that formed as a result of cell fragmentation in the process of programmed cell death (apoptosis). Among EVs, apoptotic bodies are the largest vesicles, with a size ranging from approximately 500 to 4000 nm and a density between 1.16–1.28 g/mL9,15,16. Apoptotic bodies are characterized by the presence of DNA fragments and histones along with proteins as the vesicular cargo. Apoptosis initiates with condensation of the chromatin, followed by plasma membrane blebbing and encompassing of the cellular content into distinct membrane-enclosed vesicles known as apoptotic bodies or apoptosomes. Finally, these bodies are released into the extracellular space. The outer leaflets of the lipid bilayer of apoptotic bodies contain phosphatidylserine, similar to the membrane of exosomes and MVs. These translocated phosphatidylserines can bind to Annexin V, which is recognized by phagocytes to undergo phagocytosis17. Oxidation of surface molecules causes membrane alteration that triggers binding of thrombospondin18 or the complement protein C3b, which is in turn recognized by phagocyte receptors19,20,21. Therefore, Annexin V, thrombospondin, and C3b, serve as accepted markers of apoptotic bodies21. Unlike the other two EV types, apoptotic bodies are only released during programmed cell death.
Biogenesis and composition of exosomes
Biogenesis
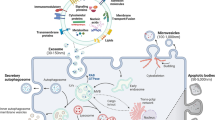
Among the three main classes of EVs, this review focuses on exosomes. EVs are cell-derived vesicles, enclosed by a lipid bilayer, with varied diameter depending on their origin. In contrast to microvesicles, which are generated by budding from the plasma membrane, exosomes are derived from the endolysosomal pathway (Figure 1). Microvesicles are closer than exosomes to the parent cells in their membrane composition, but exosomes typically contain some additional defined components3. Exosomes are formed intracellularly via endocytic invagination within the endosomal network. Endosomes at their early stage fuse with endocytic vesicles and incorporate their content into vesicles destined for recycling, degradation, or exocytosis. Early endosomes then undergo a sequence of alterations to become late endosomes or multivesicular bodies (MVBs), which are characterized by the presence of multiple small interluminal vesicles (ILVs)22. When MVBs undergo maturation, the “cargo” within them is sorted, and they then fuse with lysosomes for lysosomal degradation or fuse with the plasma membrane to be released as exosomes in the extracellular space. During this process, transmembrane proteins are incorporated into the invaginating membrane, maintaining a topological orientation similar to that of the plasma membrane1.

Biogenesis and release of extracellular vesicles: represented diagram depicts a typical extracellular vesicle biogenesis and release.
On the other hand, the biogenesis of microvesicles is distinct from exosome biogenesis. Microvesicle formation is initiated through direct outward budding followed by a fission that resembles the abscission step in cytokinesis. These vesicles, also called ectosomes23, are usually larger than exosomes, and their sizes overlap. There is an asymmetric distribution of phospholipids within the plasma membrane that is tightly regulated by aminophospholipid translocases such as flippases and floppases24,25,26. Flippases help to transfer phospholipids from the outer leaflet to the inner leaflet. Membrane budding and vesicle formation is induced by translocation of phosphatidylserine to the outer-membrane leaflet. This process is accomplished by contraction of cytoskeletal protein actin–myosin interactions27.
Composition of EVs
EVs contain signaling proteins, transcriptional regulators, nucleic acids and lipids28. The composition is based on the components of the parent cells28. For example, EVs derived from lung epithelial cells contain lung surfactant proteins29. The presence of these cell-specific proteins can act as markers to identify the origin of EVs. Moreover, the presence of certain biomarkers in body fluids also helps to identify particular disease types and the severity of the prognosis of the disease state. For example, melanoma patients have higher levels of CD63 and CAV1 in plasma exosomes compared with healthy individuals30. Furthermore, isolating exosomes from body fluid in order to engineer them using lectins or antibodies against exosome markers such as CD63, CD81, EpCAM or Rab5 is possible31. Aminopeptidase N and nebulin are two abundant membrane-associated proteins found in the MVs of nephrotic urine after ultracentrifugation32.
Compositions of exosomes
To understand the cellular composition of exosomes, a wide variety of techniques has been used, including trypsin digestion and mass spectrometry, Western blotting, and fluorescence-activated cell sorting (FACS) analyses in various types of cells. These techniques have identified a subset of cellular proteins that specifically target exosomes. Strikingly, a study involving humans and mice found that 80% of proteins from exosomes derived from dendritic cells (DCs) are conserved in both33. The protein composition of the exosomes is still mostly unknown. Due to their endosomal origin, all exosomes contain membrane transport and fusion proteins (GTPases, Annexins, and flotillin), tetraspannins (CD9, CD63, CD81, and CD82), heat shock proteins (Hsc70, Hsp 90, Hsp60 and Hsp20)34,35, proteins involved in multivesicular body biogenesis (Alix, TSG101), lipid-related proteins and phospholipases (Figure 2). Exosomes released by APCs are rich in antigen presenting proteins such as MHC class I and class II2. Exosomes derived from DCs contain CD86, an important co-stimulatory molecule for T-cells. It has been shown that different α- and β-chains of integrins, ICAM1/CD54, A33 antigen and P-selectin, or cell-surface peptidases (CD26 and CD13) are also present in exosomes33. Other than the characteristic morphology, the protein and lipid composition of exosomes is unique, providing an additional tool for their identification. Furthermore, exosomes are unique in enrichment of cholesterol, ceramide or other sphingolipids, and phospoglycerides with long and saturated fatty-acyl chains36. A recent study has shown that exosomes from mast and dendritic cells have increased levels of phosphatidylethanolamines, which have a higher rate of flipping between the two leaflets of the exosome bilayer than in cellular membranes37. There are findings that show that exosomes play a role in delivering prostaglandins to the target cells37. The outer surface of exosomes also contains saccharide groups, which are enriched in mannose, polylactosamine, α-2,6 sialic acid, and complex N-linked glycans38.

Proposed structure of a mid-size exosome (40–60 nm in diameter). A vesicle contains cytosol of the parent cell and expresses the extracellular domains of different transmembrane proteins reflecting the type of parent cell from which it is derived.
Exosomes have been reported to contain significant amounts of miRNA, other non-coding RNAs, and mRNA, which can all be transferred between cells and modulate gene expression in recipient cells39. A few studies have indicated that the RNA content in exosomes differs significantly from that of its parental cell40. Similarly, circulating EVs are carrier vehicles for a large number of miRNAs involved in cardiovascular disorders. miRNA packaging in EVs is different from the parent cell and is largely influenced by external stimuli. EVs treated with miR-150 mimic increase endothelial cell migration. miR-126 is elevated in various types of EVs and promotes re-endothelialization in vivo and therefore serves as an important regulator of angiogenesis and vascular integrity32. Another study on atherosclerotic mice injected with miR-143/145-containing EVs showed a reduction in atherosclerotic lesion formation41. Furthermore, fibroblast-derived exosomes that carry miR-21 act as a paracrine signaling factor in cardiac hypertrophy42.
Functions of exosomes
Biological function
There are many studies that have shown that exosomes derived from antigen presenting cells, such as DCs, can express major histocompatibility complex (MHC) class I and II molecules on the cell surface, which helps to induce specific immune responses by activating immune cells, such as CD8+ and CD4+ T-cells43. EVs regulate normal biological processes in a pleiotropic fashion, either directly activating cell-surface receptors of neighboring cells or merging into the plasma membrane of neighboring cells and delivering its cargos, including transcription factors, oncogenes, miRNAs, mRNAs and infectious particles44,45,46. The exosomes thus serve as important effector molecules that modulate normal physiological functions of the body, such as stem cell maintenance45, tissue repair47, immune surveillance48 and blood coagulation49.
EVs either activate or inhibit the function of regulatory T cells; suppress natural killer cells (NKs) and CD8+ cell activity; or activate monocytes, B cells and NK cells3. EVs have an intrinsic adjuvant effect, which enables them to become efficient immune modulatory molecules that transfer antigens between APCs. EVs isolated from mast cells contain relatively high HSP60 and HSC70 content that promotes DC maturation in mice2. In addition, bacteria-infected macrophages release EVs carrying microbial antigens and pathogen-associated molecular patterns that promote an inflammatory response by macrophages in a TLR-dependent manner50.
Pathological function
Role of exosomes in cardiac disease
Exosomes are known to be involved in many cardiovascular physiological and pathological disorders, such as cardiomyocyte hypertrophy, peripartum cardiomyopathy and sepsis-induced cardiomyopathy. In response to cardiac stresses, such as myocardial infarction, cardiac valve disease, and systemic hypertension, the heart undergoes extensive cardiac remodeling that results in cardiac fibrosis and pathological growth of cardiomyocytes or hypertrophy51,52. In the hypertrophic heart, cardiac fibroblasts induce and modify cardiomyocyte hypertrophy by secreting different growth factors and extracellular matrix components53. Recently, a study by Bang et al showed that cardiac fibroblasts secrete exosomes enriched in miRNAs, which are often degraded intracellularly. Approximately 25.5% of fibroblast-derived exosomes contain these “star” miRNAs. miRNA profiling of exosomes revealed that exosomal miR-21* is a potent paracrine-like miRNA molecule that induces cardiomyocyte hypertrophy. This can be mediated by silencing SORBS2 (sarcoplasmic protein sorbin and SH3 domain-containing protein 2) and PDLIM5 (PDZ and LIM domain 5) proteins52.
Peripartum/postpartum cardiomyopathy (PPCM) is a critical, potentially life-threatening pregnancy-associated cardiomyopathy characterized by sudden heart failure during the last month of pregnancy and/or in the first few months postpartum51. Cathepsin D is cleaved from a 16-kDa N-terminal prolactin fragment (16K PRL) in the full-length nursing hormone prolactin (PRL) and is believed to be a potential factor in initiating PPCM54. Although the underlying molecular mechanisms are not clear, Halkein et al reported that 16K PRL not only induced the expression of miR-146a in endothelial cells (ECs) but also enhanced the release of miR-146a-enriched exosomes from ECs. These EC-derived exosomes were absorbed by cardiomyocytes, resulting in elevation of miR-146a levels. Consequently, the expression of Erbb4, Notch1, and Irak1 was decreased in cardiomyocytes, which ultimately led to impaired metabolic activity and contractile function51,55. Furthermore, levels of exosomal miR-146a were found to be significantly higher in plasma from patients with acute PPCM than healthy postpartum controls and patients with dilated cardiomyopathy. Therefore, exosomal miR-146a may also serve as a highly specific blood biomarker that is useful for diagnosis of patients with PPCM.
Early during diabetes, high glucose levels in the bloodstream can lead to endothelial dysfunction. This promotes abnormal vascular growth that triggers the progression of atherosclerosis in patients with diabetes mellitus56. In a recent report, Wang et al showed that in response to hyperglycemia, exosomes derived from cardiomyocytes harbor a variety of miRNAs, which may be transferred to adjacent ECs and modulate their function57. Furthermore, their findings revealed that cardiomyocyte-derived exosomes from diabetic type 2 diabetic Goto-Kakizaki GK rats contain higher levels of miR-320 and lower levels of miR-126 and Hsp20 proteins than exosomes collected from healthy cardiomyocytes. Importantly, cardiomyocyte-derived exosomal miR-320 can be transferred to ECs and consequently down-regulates the expression of IGF-1, Hsp20, and Ets-2, leading to inhibition of EC proliferation, migration and tube formation. It has also been shown by Wang et al that diabetic cardiomyocytes release exosomes containing lower levels of Hsp20 than exosomes from normal cardiomyocytes, which is responsible for hyperglycemia-induced cell death. Overexpression of Hsp20 significantly reduced cardiac dysfunction, hypertrophy, apoptosis, fibrosis, and microvascular rarefaction in diabetic mice. Overall, this study uncovered a novel mechanism underlying the impairment of myocardial angiogenesis in diabetes, which may be caused by the secretion of anti-angiogenic exosomes from cardiomyocytes58.
Myocardial dysfunction is one of the main predictors of poor outcome in septic patients, with mortality rates of approximately 70%59. Evidence suggests that platelet-derived exosomes might be involved in myocardial dysfunction in sepsis patients60. Numerous studies have indicated that the presence of exosomes in the plasma of sepsis patients may cause vascular and cardiac dysfunction in sepsis51. Studies have shown that lipopolysaccharide (LPS) triggers increased generation of nitric oxide (NO) and subsequently induces the release of exosomes from platelets during sepsis. These LPS-induced platelet-derived exosomes contain higher levels of NADPH oxidase, nitric oxide synthases (NOS) and protein disulfide isomerase (PDI) than healthy exosomes. Furthermore, it has been shown that NO-induced and human septic platelet-derived exosomes induce caspase-3 activation and apoptosis of target ECs through generation of active ROS/RNS by NADPH oxidase and NO synthase type II61. In addition, one report provided evidence that circulating platelet-derived exosomes from septic patients induced myocardial dysfunction in isolated heart and papillary muscle preparations. This effect was further enhanced by in vivo pre-exposure of exosomes to LPS60.
Role of exosomes in Neurodegenerative disease
Neurodegenerative disorders are characterized by the deposition of misfolded, aggregated forms of specific proteins in defined neuroanatomical locations. Recent research has revealed that the majority of proteins involved in neurodegenerative diseases are transported in exosomes. Exosomes containing aggregation-prone proteins involved in Parkinson's disease (PD), Alzheimer's disease (AD), Creutzfeldt-Jakob disease (CJD), and amyotrophic lateral sclerosis (ALS) have all been found in the cerebral spinal fluid and blood of patients affected by these disorders. Moreover, exosomes provide a supportive environment that can induce conformational change of native proteins into aggregates that can be transmitted to aggregate-free cells in the brain62. In Parkinson's disease, mutated α-synuclein proteins form intracellular oligomers (known as Lewy's bodies) that can be secreted via exosomes to the extracellular milieu and taken up by nearby cells, thus spreading the disease from cell to cell within the brain63. Further investigation on this subject by Danzer et al has shown that autophagy acts as a protective mechanism in cells and constitutes a major degradation pathway for α-synuclein oligomers64. Any deregulation of neuronal autophagy might promote the aggregation of these proteins and their secretion by exosome release, thereby spreading and causing neurodegeneration63.
In Alzheimer's disease, tau protein aggregates form filamentous intracellular inclusions that can spread from affected nerve and glial cells to healthy cells, thus spreading the disease. Studies have shown that extracellular α-amyloid aggregates can induce tau pathology in transgenic mice and finally promote neurodegeneration65,66. Furthermore, exosome-associated tau and α-amyloid have been proposed to be present on the exosomal surface, which can act as a platform for β-amyloid aggregation after protein conformational modifications63,67. Prions are abnormally folded proteins with the ability to propagate in the central nervous system, causing fatal neurodegenerative disorders such as Creutzfeldt–Jakob disease and Gerstmann–Sträussler–Scheinker syndrome. Infectious prion proteins (PrPSc) have been identified in exosomes derived from the conditioned media of mammalian neurons68,69. These exosomes were internalized by normal cells and transformed naturally occurring cellular prion proteins (PrPc) into misfolded infectious prion proteins (PrPSc) and thus spread the disease63. Amyotrophic lateral sclerosis (ALS) is a neurodegenerative disorder of motor neurons. Mutations in superoxide dismutase 1 (SOD1) and TDP-43 are involved in inherited forms of ALS, and both these proteins have exhibited template-directed induction of pathological misfolding. Furthermore, these proteins have been found to be associated with exosomes, suggesting a potential role of extracellular vesicles in intercellular transfer of misfolded SOD1 and TDP4362,70.
Role of exosomes in cancer
Direct interaction between tumor cells and their environment is absolutely necessary for cancer progression. Growing evidence indicates that exosomes from cancer cells transfer oncogenic proteins and nucleic acids that modulate the activity of recipient cells and play decisive roles in tumorigenesis, cell proliferation, progression, metastasis, and drug resistance. Prostate cancer cell-derived exosomes containing oncogenic proteins (Ras superfamily GTPases), mRNA (H-ras and K-ras), and miRNAs (miR-125b, miR-130b, and miR-155) can induce neoplastic transformation of adipose-derived stem cells (ASCs)71,72. Tumor-derived exosomes can also activate endothelial cells to support tumor angiogenesis and thrombosis. Exosomes derived from hypoxic glioblastoma cells are potent inducers of angiogenesis71. K562 leukemia cell-derived exosomal miR-92a targets integrin α5 to enhance endothelial cell migration and tube formation73. Hypoxic K562 cells released miR-210-enriched exosomes that promote the angiogenic activity of endothelial cells74. Exosomes also contribute to creating an immunosuppressive microenvironment by inducing apoptosis and impairing the function of effector T cells and natural killer cells (NKs), inhibiting dendritic cell (DC) differentiation, expanding myeloid-derived suppressor cells (MDSCs), and promoting regulatory T cell (Treg) activity. Exosomes derived from tumors can also convert fibroblasts and mesenchymal stromal cells into myofibroblasts to facilitate tumor angiogenesis and metastasis. Tumor-derived exosomes can mobilize neutrophils and skew M2 polarization of macrophages to promote tumor progression. Moreover, tumor-derived exosomes can help tumor cells develop drug resistance by transferring multidrug-resistant proteins and miRNAs, exporting anti-cancer drugs, and neutralizing antibody-based drugs. In turn, exosomes from activated T cells, macrophages, and stromal cells can promote tumor metastasis and drug resistance71.
Role of exosomes in liver inflammation
Studies have shown that exosomes are also involved in liver inflammation. It has been revealed that repeated injection of exosomes isolated from peripheral blood of mice fed a high-fat diet to mice fed a regular diet resulted in activated immature CD11b+Ly6ChiLy6G− myeloid cell accumulation in the liver that caused chronic inflammation and promoted obesity-related disorders, such as fatty liver disease75,76. These studies clearly indicate the pathological role of exosomes in metabolic liver disease. Electron microscope images showed that exosome number is increased in the cells after treatment with LPS compared with the normal unstimulated cells (Figure 3). This suggests that exosomes may make a significant contribution during inflammation-based modulation of cells.

Electron micrographs of mouse bone marrow derived primary macrophage cells treated with or without lipopolysaccharide (LPS). (A) Exosomes in the untreated cells; (B) exosomes after treatment with LPS; (C) exosomes secreted outside the cells; MVB, multivesicular body; Bars: 100 nm.
Different strategies to attenuate adverse functions of exosomes during disease
After understanding extracellular vesicle-mediated disease pathogenesis, it is important to understand the ways that they could be inhibited. Several approaches could be used to attenuate extracellular vesicle function and inhibit the development of diseases. These include their formation, release, cell uptake or targeting extracellular vesicle components that are responsible for disease pathogenesis.
Targeting formation
Different cellular components are recognized to be critical for extracellular vesicle formation. The best approach to inhibit their formation is inhibiting cellular components that aid in formation of exosomes by small molecules or RNAi. Therefore, using inhibitors of sphingomyelinase or amiloride, a blood-pressure-lowering drug, can inhibit ceramide formation and thereby vesicle formation3. The use of amiloride was also found to be effective in reducing mouse and human tumor cell growth by blocking EVs harboring membrane-associated heat shock protein 72 (HSP72)77. Inhibiting the interaction of syndecan proteoglycans and their cytoplasmic adaptor syntenin with the exosomal protein ALIX could attenuate exosome release. Blocking specific tetraspanins, eg, TSPAN30/CD63, which are important for extracellular vesicle formation, could be another alternative approach to inhibit EVs by reducing the formation tumors3.
Targeting release
Many proteins have been found to be important for the release of EVs, including exosomes, but the detailed mechanism of vesicle release remains unclear and is likely to vary among cell types. One approach is to inhibit their release into the extracellular space so that their effect on other cells is prevented. For example, EV formation and release into the extracellular space relies on the endosomal sorting complex required for transport (ESCRT) and/or ceramide and sphingosine 1-phosphate. Targeting these complexes with small molecules or RNAi inhibits EV release into the extracellular space. The GTPase RAB27A73, another molecule involved in exosome release in some tumor cells, could also act as a therapeutic target for reducing tumor growth78. Furthermore, using GW4869 and specific small interfering RNA blocks the biosynthesis of ceramide and thereby inhibits exosome release from the affected cells to the neighboring non-transformed cells. This could be another approach to control intracellular communications mediated by exosomes. This approach has been shown to be effective for reducing sepsis-induced inflammatory responses and improving cardiac dysfunction, as well as lowering the presence of circulating miRNAs in blood cancer patients79,80.
Targeting uptake
Inhibiting extracellular vesicle uptake would be another approach to inhibit disease-causing EVs. There are many mechanisms for uptake of extracellular vesicles, but the detailed mechanisms in extracellular vesicle trafficking and targeting are not as well understood. Phosphatidylserine (PS) is an important surface molecule, and therefore, blocking PS with diannexin can reduce adhesion of extracellular vesicles released from tumor cells to non-tumor cells81,82. Although this strategy had some effect on reducing tumors in a mouse model, there are also some disadvantages3. EV uptake is also mediated by clathrin-dependent endocytosis and caveolin-mediated (clathrin-independent) uptake, macropinocytosis, phagocytosis, and lipid raft-mediated internalization. It is also important to further look into these cellular processes to inhibit the uptake of EVs and thereby minimize disease progression83.
Targeting trafficking
Blocking specific signaling molecules responsible for the trafficking of EVs might be another approach to inhibit their activity. For example, in a melanoma model, it has been shown that monoclonal antibodies for FASL could target FASL1 present on the cell surface of EVs and thus reduce tumor development. However, one of the great limitations of this process is the specificity, which affects the immune system globally3,84. Recent studies have shown that there are specific cofactors for nuclear exosomes that aid in exosome RNA synthesis and stability. For example, nucleoplasm-specific Nuclear EXosome Targeting (NEXT) complex constitutes a non-nucleolar hMTR4-containing complex that targets PROMoter uPstream Transcripts (PROMPTs) for rapid exosomal turnover in humans85. Therefore, targeting these complexes could be an approach to inhibit overall exosome turnover.
Potential role of exosomes and microvesicles in therapy
The natural cargo carrying property of EVs makes them a promising therapeutic agent for different disease states, including cancer and degenerative diseases.
Stem cell-derived exosomes and microvesicles have a protective effect
For a long time, mesenchymal stem cells (MSCs) have been used for preclinical and clinical studies, including acute myocardial infarction, stroke, acute kidney failure, and many others (http://clinicaltrials.gov). Recent studies have found that lung-trapped MSCs secrete an anti-inflammatory protein, TSG-6, which plays a beneficial role in myocardial infarction86. Another study that explored the use of MSC-derived exosomes after myocardial ischemia in mice observed a reduction in infarct size87. Taken together, these studies suggest that exosomes play an important role in regenerative therapies88. In a report on middle cerebral artery occlusions in a stroke model, mice were treated with MSC-derived exosomes, which were known to contribute to increased neurite branch numbers as well as total neurite length after stroke89. It has been shown that human umbilical cord MSC-derived exosomes promote proliferation and inhibit apoptosis of skin cells in rat burn models. Moreover, the wounds treated with MSC-derived exosomes exhibited accelerated re-epithelialization with increased expression of CK19, PCNA, and collagen I in vivo90. MSC-derived exosomes can potentially play regenerative roles in tendinopathy by developing exosomes as a non-surgical and non-cellular treatment option for tendon repair (Tetta C 2012). Recent studies have shown that cardiac cell-derived exosomes mediate protective cardiac functions in a murine model of myocardial infarction91,92. Exosomes derived from MSCs are currently being used for fracture healing where they might exert an important regulatory function in osteogenic differentiation of bone marrow stem cells. These effects are mediated by upregulation of miRNAs, such as miR-21, miR-125b and miR-445493. Another study has shown that mean pulmonary artery pressure (mPAP) and mean right ventricle pressure (mRVP) were reduced significantly in pulmonary arterial hypertensive rats who received intravenous injection of MSC-derived MVs or MSCs94. More recently, it has been shown that in diabetes the circulating MV and endothelial progenitor cell-derived MV levels are lower due to a decreased level of miR-126, and therefore, this could be used as a potential target to treat patients with diabetic vascular complications95.
Pharmacological delivery systems for molecules of interest
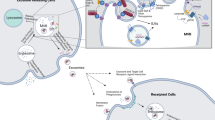
Exosomes can serve as an efficient carrier system to deliver different pharmacological molecules to target cells or tissues. These molecules can be further modified and reinserted into exosomes for different therapeutic applications (Figure 4).

Exosomes play roles in drug delivery: exosomes isolated from different cell types are rich in miRNA, RNA and protein. These molecules can further modified and reinserted into the exosomes for different therapeutic applications.
Advantages of exosomes as carrier vehicles
Liposomes and polymeric nanoparticles are two exciting drug delivery platforms that hold great promise. However, the major concern with liposome-mediated delivery is the stability, toxicity and ability to evade the host immune system96. Conversely, polymeric nanoparticles have better stability, but their toxicity and rapid clearance by the immune system is a concern97,98. In this respect, exosomes or exosome mimetics are more advantageous for an ideal drug delivery system with more of the desirable features. Exosomes exhibit an increased circulating half-life that enables them to travel long distances within the body under both physiological and pathological conditions99. Additionally, exosomes have more biocompatibility, which makes them suitable to host soluble drugs at room temperature (RT). Drugs such as the low molecular weight antioxidant curcumin100, anticancer agents such as Doxorubicin (Dox)101 and Paclitaxel (PTX)102, and Rhodamine-123102 can be incorporated within exosomes. However, the fact that exosomes are already packed with intrinsic natural molecules presents a problem, and thus, it is often hard to load additional molecules into them103. Because they are such small molecules and they carry cell surface molecules, exosomes have a high affinity for tissues104,105,106, and they possess a natural targeting capacity and thus have less off-target effects99, which makes them a better option for drug-delivery systems. Moreover, exosomes have been shown to cross the blood-brain barrier and selectively be taken up by brain microglial cells, and therefore, they can be used in the treatment of diseases of the central nervous system107. Furthermore, the immunogenicity of exosomes is very low compared to liposomes and virus-based drug delivery systems106.
Manipulation of exosomes is relatively simple and can be performed by isolating them from cell cultures or a patient's body fluids, serum, and plasma, and after modification, they can be subsequently transferred back to the same patient105. In addition, exosome mimetics can be created synthetically by liposomes harboring only essential components of natural exosomes99. Phase I clinical trials have already been performed with dendritic cell-derived exosomes as cell-free cancer vaccines108. In other clinical trial for patients with stage III/IV metastatic melanoma, dendritic cells (DC) were collected and pulsed with MAGE 3 peptides to be used by MHC for antigen presentation. Next, autologous EVs were reintroduced into patients to evoke an immune response against melanoma. Mostly, patients tolerated EV administration for up to 21 months with mild inflammatory responses109. Another trial for patients with non-small cell lung cancer was performed to assess the safety, feasibility, and efficacy of DC-derived exosomes incubated with the MAGE tumor antigens weekly for 4 weeks, which manifested low-level inflammatory responses110. These results encouraged testing of DC-derived EVs for non-small cell lung cancer treatment in a Phase II clinical trial (http://clinicaltrials.gov/show/NCT01159288). EVs are also being tested as a curcumin delivery agent for colon cancer patients (http://clinicaltrials.gov/show/NCT01294072).
Extracellular vesicles as gene therapy devices
Accumulating evidence has revealed that small interfering RNAs can be utilized as therapeutic agents, particularly for cancer and viral infections111. However, due to the lack of appropriate delivery systems, their clinical application is still circumscribed. Certain intrinsic properties of exosomes, such as their small size and natural ability to carry miRNA, their cellular membranes with multiple adhesive proteins on the surface and their ability to cross the blood–brain barrier, make exosomes an excellent gene delivery system for gene therapy104,112,113.
Ohno et al modified human embryonic kidney cell line 293 (HEK293) cells with forced expression of GE11 or EGF using a pDisplay expression vector. They then isolated exosomes from culture supernatants that carried GE11 or EGF on their surface. Finally, they injected these modified HEK293 cell-derived exosomes to EGFR-expressing cancer tissues114. Akao et al successfully entrapped chemically modified miR-143BP in microvesciles and transfected THP-1 macrophages with the same RNA molecules, followed by collection of exosomes by ultracentrifugation. The exosomes from these modified THP-1 macrophages were shown to effectively target tumors and the kidneys and treat cancer and other diseases115.
Exogenous siRNA could also be loaded into exosomes by electroporation. In a model of Alzheimer's disease, Erviti et al loaded GAPDH siRNA into DC-derived exosomes to specifically deliver them to neurons, microglia, and oligodendrocytes in the brain. They also showed that exosome-mediated delivery of siRNA against the BACE1 gene resulted in strong mRNA (60%) and protein (62%) knockdown of BACE1 in wild-type mice104. This group thus demonstrated that exosome-mediated siRNA delivery is a promising therapeutic agent against neurodegenerative diseases.
Protein cargo
The size and structural proximity of exosomes with cellular components make them easy to use as a specific drug delivery medium in which the drugs are usually protein molecules. A study has shown that exosomes can cross the blood-brain barrier and improve PD status when loaded with the antioxidant protein catalase ex vivo, which makes the use of exosomes a promising option for PD therapy116. CD44 expression is high in tumorigenic and metastatic hepatocellular cancer stem cells (CSCs). Therefore, anti-CD44 antibody-coated liposomes that can deliver doxorubicin directly to these cells would have great potential for targeted therapy in cancer117. Recently, work by Meyer et al has shown that vesicular stomatitis virus glycoprotein (VSVG) can both load protein cargo onto exosomes and increase their intracellular therapeutic protein delivery ability via exosome-based vehicles and thus shows promise for exosome-based delivery118.
Challenges and future direction
Using exosomes as drug delivery vehicles has several advantages over nano-based or liposome-based therapies. Particularly, exosomes can avoid phagocytosis or degradation by macrophages and can also circulate for prolonged times; however, there are a number of limitations and challenges for translation into clinical therapies. A key problem prohibiting exploring exosomes in clinical applications is the availability of pure exosomes from the body119. Because of the low number of exosomes, purifying them is also difficult120. Each of the available methods of exosome isolation from cell culture supernatants or different biological fluids, including milk, urine, plasma, amniotic fluid, saliva, and cerebrospinal fluid121 have different advantages and disadvantages. It is important to determine the best source from which more pure, well-characterized exosomes with high quality can be obtained122. Characterization of exosomes using different available techniques such as electron microscopy, FACS, and Western blot analyses have limitations and therefore cannot be used as independent methods to characterize the biophysical and biochemical properties of exosomes. There should be definite and reproducible techniques to characterize MVs clinically and use them as attractive biomarkers for different pathological conditions44.
There are controversies regarding how to define EV dosage, number of vesicle particles, the amount of vesicle protein, or expressing dosage as a vesicle number to protein ratio. This optimization is important to define EV dosage for clinical trials28. Next, enhancing efficient loading of various cargos and targeting proficiencies of exosomes without altering the structure and content of exosomal membranes is important for increasing the use of exosomes while maintaining their functional efficacy. It has also been found that combining exosomes with different therapeutic cargoes often makes them immunogenic based on the nature of parental donor cells, and therefore, further studies are needed to delineate immunogenic reactions after administration. Ongoing research seeks to find ways of developing and standardizing appropriate methods to modify exosome contents in a loading process. Further exploration of the structure and function of exosomes will contribute to the clinical application of exosomes.
Conclusion
Exosomes have emerged as an important drug delivery vehicle because they can be loaded with a variety of cargo molecules that can transfer bioactive proteins, lipids, and nucleic acids. Due to their size and inherent similarity to the parent cell, they can be exploited for therapy in different clinical perspectives. The paracrine effect of exosomes and prolonged retention in the circulation make them increasingly important. Therefore, exosomes act as naturally occurring nanoparticles, and efforts will be made to better understand their properties and function.
Abbreviations
EV, extracellular vesicles; AB, apoptotic bodies; MV, microvesicles; PS, phosphatidylserine; ILV, interluminal vesicles; MT1, membrane type 1; VAMP3, vesicle-associated membrane protein 3; MHC, major histocompatibility complex; NK, natural killer cells; PPCM, peripartum/postpartum cardiomyopathy; PRL, prolactin; EC, endothelial cell; PD, Parkinson's disease; AD, Alzheimer's disease; CJD, Creutzfeldt-Jakob disease; ALS, amyotrophic lateral sclerosis; PrPc, prion proteins; SOD1, superoxide dismutase 1; ASC, adipose-derived stem cell; HSP72, heat shock protein 72; ESCRT, endosomal sorting complex required for transport; ICAM1, intercellular adhesion molecule 1; CSC, cancer stem cell; VSVG, vesicular stomatitis virus glycoprotein.
References
Muralidharan-Chari V, Clancy JW, Sedgwick A, D'Souza-Schorey C . Microvesicles: mediators of extracellular communication during cancer progression. J Cell Sci 2010; 123: 1603–11.
Robbins PD, Morelli AE . Regulation of immune responses by extracellular vesicles. Nat Rev Immunol 2014; 14: 195–208.
El Andaloussi S, Mager I, Breakefield XO, Wood MJ . Extracellular vesicles: biology and emerging therapeutic opportunities. Nat Rev Drug Discov 2013; 12: 347–57.
Yanez-Mo M, Siljander PR, Andreu Z, Zavec AB, Borras FE, Buzas EI, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles 2015; 4: 27066.
Samanta S, Balasubramanian S, Rajasingh S, Patel U, Dhanasekaran A, Dawn B, et al. MicroRNA: a new therapeutic strategy for cardiovascular diseases. Trends Cardiovasc Med 2016; 26: 407–19.
Balasubramanian S, Rajasingh S, Thangavel J, Dawn B, Rajasingh J . Chapter 3 - Exosome function in miRNA-mediated paracrine effects. Mesenchymal stem cell derived exosomes. Boston: Academic Press; 2015. p 37–62.
Yin M, Loyer X, Boulanger CM . Extracellular vesicles as new pharmacological targets to treat atherosclerosis. Eur J Pharmacol 2015; 763: 90–103.
Nomura S . Extracellular vesicles and blood diseases. Int J Hematol 2017; 105: 392–405.
Revenfeld AL, Baek R, Nielsen MH, Stensballe A, Varming K, Jorgensen M . Diagnostic and prognostic potential of extracellular vesicles in peripheral blood. Clin Ther 2014; 36: 830–46.
Kowal J, Arras G, Colombo M, Jouve M, Morath JP, Primdal-Bengtson B, et al. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc Natl Acad Sci U S A 2016; 113: E968–77.
Kalluri R . The biology and function of exosomes in cancer. J Clin Invest 2016; 126: 1208–15.
Giusti I, D'Ascenzo S, Dolo V . Microvesicles as potential ovarian cancer biomarkers. Biomed Res Int 2013; 2013: 703048.
Badimon L, Suades R, Fuentes E, Palomo I, Padro T . Role of platelet-derived microvesicles as crosstalk mediators in atherothrombosis and future pharmacology targets: a link between inflammation. Atheroscler Thromb Front Pharmacol 2016; 7: 293.
Pugholm LH, Revenfeld AL, Sondergaard EK, Jorgensen MM . Antibody-based assays for phenotyping of extracellular vesicles. Biomed Res Int 2015; 2015: 524817.
Kerr JF, Wyllie AH, Currie AR . Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br J Cancer 1972; 26: 239–57.
New SE, Aikawa E . Role of extracellular vesicles in de novo mineralization: an additional novel mechanism of cardiovascular calcification. Arterioscler Thromb Vasc Biol 2013; 33: 1753–8.
Martinez MC, Freyssinet JM . Deciphering the plasma membrane hallmarks of apoptotic cells: phosphatidylserine transverse redistribution and calcium entry. BMC Cell Biol 2001; 2: 20.
Friedl P, Vischer P, Freyberg MA . The role of thrombospondin-1 in apoptosis. Cell Mol Life Sci 2002; 59: 1347–57.
Takizawa F, Tsuji S, Nagasawa S . Enhancement of macrophage phagocytosis upon iC3b deposition on apoptotic cells. FEBS Lett 1996; 397: 269–72.
Erwig LP, Henson PM . Clearance of apoptotic cells by phagocytes. Cell Death Differ 2008; 15: 243–50.
Savill J, Hogg N, Ren Y, Haslett C . Thrombospondin cooperates with CD36 and the vitronectin receptor in macrophage recognition of neutrophils undergoing apoptosis. J Clin Invest 1992; 90: 1513–22.
Akers JC, Gonda D, Kim R, Carter BS, Chen CC . Biogenesis of extracellular vesicles (EV): exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J Neurooncol 2013; 113: 1–11.
Hess C, Sadallah S, Hefti A, Landmann R, Schifferli JA . Ectosomes released by human neutrophils are specialized functional units. J Immunol 1999; 163: 4564–73.
Zwaal RF, Schroit AJ . Pathophysiologic implications of membrane phospholipid asymmetry in blood cells. Blood 1997; 89: 1121–32.
Bevers EM, Comfurius P, Dekkers DW, Zwaal RF . Lipid translocation across the plasma membrane of mammalian cells. Biochim Biophys Acta 1999; 1439: 317–30.
Leventis PA, Grinstein S . The distribution and function of phosphatidylserine in cellular membranes. Annu Rev Biophys 2010; 39: 407–27.
McConnell RE, Higginbotham JN, Shifrin DA Jr, Tabb DL, Coffey RJ, Tyska MJ . The enterocyte microvillus is a vesicle-generating organelle. J Cell Biol 2009; 185: 1285–98.
Xu R, Greening DW, Zhu HJ, Takahashi N, Simpson RJ . Extracellular vesicle isolation and characterization: toward clinical application. J Clin Invest 2016; 126: 1152–62.
Moon HG, Cao Y, Yang J, Lee JH, Choi HS, Jin Y . Lung epithelial cell-derived extracellular vesicles activate macrophage-mediated inflammatory responses via ROCK1 pathway. Cell Death Dis 2015; 6: e2016.
Properzi F, Logozzi M, Fais S . Exosomes: the future of biomarkers in medicine. Biomark Med 2013; 7: 769–78.
Lin J, Li J, Huang B, Liu J, Chen X, Chen XM, et al. Exosomes: novel biomarkers for clinical diagnosis. ScientificWorldJournal 2015; 2015: 657086.
Rood IM, Deegens JK, Merchant ML, Tamboer WP, Wilkey DW, Wetzels JF, et al. Comparison of three methods for isolation of urinary microvesicles to identify biomarkers of nephrotic syndrome. Kidney Int 2010; 78: 810–6.
Thery C, Zitvogel L, Amigorena S . Exosomes: composition, biogenesis and function. Nat Rev Immunol 2002; 2: 569–79.
Gupta S, Knowlton AA . HSP60 trafficking in adult cardiac myocytes: role of the exosomal pathway. Am J Physiol Heart Circ Physiol 2007; 292: H3052–6.
Zhang X, Wang X, Zhu H, Kranias EG, Tang Y, Peng T, et al. Hsp20 functions as a novel cardiokine in promoting angiogenesis via activation of VEGFR2. PLoS One 2012; 7: e32765.
Trajkovic K, Hsu C, Chiantia S, Rajendran L, Wenzel D, Wieland F, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008; 319: 1244–7.
Laulagnier K, Motta C, Hamdi S, Roy S, Fauvelle F, Pageaux JF, et al. Mast cell- and dendritic cell-derived exosomes display a specific lipid composition and an unusual membrane organization. Biochem J 2004; 380: 161–71.
Batista BS, Eng WS, Pilobello KT, Hendricks-Munoz KD, Mahal LK . Identification of a conserved glycan signature for microvesicles. J Proteome Res 2011; 10: 4624–33.
Pegtel DM, Cosmopoulos K, Thorley-Lawson DA, van Eijndhoven MA, Hopmans ES, Lindenberg JL, et al. Functional delivery of viral miRNAs via exosomes. Proc Natl Acad Sci U S A 2010; 107: 6328–33.
Mittelbrunn M, Gutierrez-Vazquez C, Villarroya-Beltri C, Gonzalez S, Sanchez-Cabo F, Gonzalez MA, et al. Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nat Commun 2011; 2: 282.
Leroyer AS, Isobe H, Leseche G, Castier Y, Wassef M, Mallat Z, et al. Cellular origins and thrombogenic activity of microparticles isolated from human atherosclerotic plaques. J Am Coll Cardiol 2007; 49: 772–7.
Emanueli C, Shearn AI, Angelini GD, Sahoo S . Exosomes and exosomal miRNAs in cardiovascular protection and repair. Vascul Pharmacol 2015; 71: 24–30.
Zitvogel L, Regnault A, Lozier A, Wolfers J, Flament C, Tenza D, et al. Eradication of established murine tumors using a novel cell-free vaccine: dendritic cell-derived exosomes. Nat Med 1998; 4: 594–600.
Lee Y, El Andaloussi S, Wood MJ . Exosomes and microvesicles: extracellular vesicles for genetic information transfer and gene therapy. Hum Mol Genet 2012; 21: R125–34.
Ratajczak J, Miekus K, Kucia M, Zhang J, Reca R, Dvorak P, et al. Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: evidence for horizontal transfer of mRNA and protein delivery. Leukemia 2006; 20: 847–56.
Camussi G, Deregibus MC, Bruno S, Grange C, Fonsato V, Tetta C . Exosome/microvesicle-mediated epigenetic reprogramming of cells. Am J Cancer Res 2011; 1: 98–110.
Gatti S, Bruno S, Deregibus MC, Sordi A, Cantaluppi V, Tetta C, et al. Microvesicles derived from human adult mesenchymal stem cells protect against ischaemia-reperfusion-induced acute and chronic kidney injury. Nephrol Dial Transplant 2011; 26: 1474–83.
Raposo G, Nijman HW, Stoorvogel W, Liejendekker R, Harding CV, Melief CJ, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med 1996; 183: 1161–72.
Del Conde I, Shrimpton CN, Thiagarajan P, Lopez JA . Tissue-factor-bearing microvesicles arise from lipid rafts and fuse with activated platelets to initiate coagulation. Blood 2005; 106: 1604–11.
Bhatnagar S, Shinagawa K, Castellino FJ, Schorey JS . Exosomes released from macrophages infected with intracellular pathogens stimulate a proinflammatory response in vitro and in vivo. Blood 2007; 110: 3234–44.
Ailawadi S, Wang X, Gu H, Fan GC . Pathologic function and therapeutic potential of exosomes in cardiovascular disease. Biochim Biophys Acta 2015; 1852: 1–11.
Bang C, Batkai S, Dangwal S, Gupta SK, Foinquinos A, Holzmann A, et al. Cardiac fibroblast-derived microRNA passenger strand-enriched exosomes mediate cardiomyocyte hypertrophy. J Clin Invest 2014; 124: 2136–46.
Hilfiker-Kleiner D, Sliwa K, Drexler H . Peripartum cardiomyopathy: recent insights in its pathophysiology. Trends Cardiovasc Med 2008; 18: 173–9.
Hilfiker-Kleiner D, Kaminski K, Podewski E, Bonda T, Schaefer A, Sliwa K, et al. A cathepsin D-cleaved 16 kDa form of prolactin mediates postpartum cardiomyopathy. Cell 2007; 128: 589–600.
Halkein J, Tabruyn SP, Ricke-Hoch M, Haghikia A, Nguyen NQ, Scherr M, et al. MicroRNA-146a is a therapeutic target and biomarker for peripartum cardiomyopathy. J Clin Invest 2013; 123: 2143–54.
Nakagami H, Kaneda Y, Ogihara T, Morishita R . Endothelial dysfunction in hyperglycemia as a trigger of atherosclerosis. Curr Diabetes Rev 2005; 1: 59–63.
Wang X, Huang W, Liu G, Cai W, Millard RW, Wang Y, et al. Cardiomyocytes mediate anti-angiogenesis in type 2 diabetic rats through the exosomal transfer of miR-320 into endothelial cells. J Mol Cell Cardiol 2014; 74: 139–50.
Wang X, Gu H, Huang W, Peng J, Li Y, Yang L, et al. Hsp20-mediated activation of exosome biogenesis in cardiomyocytes improves cardiac function and angiogenesis in diabetic mice. Diabetes 2016; 65: 3111–28.
Romero-Bermejo FJ, Ruiz-Bailen M, Gil-Cebrian J, Huertos-Ranchal MJ . Sepsis-induced cardiomyopathy. Curr Cardiol Rev 2011; 7: 163–83.
Azevedo LC, Janiszewski M, Pontieri V, Pedro Mde A, Bassi E, Tucci PJ, et al. Platelet-derived exosomes from septic shock patients induce myocardial dysfunction. Crit Care 2007; 11: R120.
Gambim MH, do Carmo Ade O, Marti L, Verissimo-Filho S, Lopes LR, Janiszewski M . Platelet-derived exosomes induce endothelial cell apoptosis through peroxynitrite generation: experimental evidence for a novel mechanism of septic vascular dysfunction. Crit Care 2007; 11: R107.
Howitt J, Hill AF . Exosomes in the pathology of neurodegenerative diseases. J Biol Chem 2016; 291: 26589–97.
De Toro J, Herschlik L, Waldner C, Mongini C . Emerging roles of exosomes in normal and pathological conditions: new insights for diagnosis and therapeutic applications. Front Immunol 2015; 6: 203.
Danzer KM, Kranich LR, Ruf WP, Cagsal-Getkin O, Winslow AR, Zhu L, et al. Exosomal cell-to-cell transmission of alpha synuclein oligomers. Mol Neurodegener 2012; 7: 42.
Gotz J, Chen F, van Dorpe J, Nitsch RM . Formation of neurofibrillary tangles in P301l tau transgenic mice induced by Abeta 42 fibrils. Science 2001; 293: 1491–5.
Bolmont T, Clavaguera F, Meyer-Luehmann M, Herzig MC, Radde R, Staufenbiel M, et al. Induction of tau pathology by intracerebral infusion of amyloid-beta -containing brain extract and by amyloid-beta deposition in APP x Tau transgenic mice. Am J Pathol 2007; 171: 2012–20.
Simons M, Raposo G . Exosomes--vesicular carriers for intercellular communication. Curr Opin Cell Biol 2009; 21: 575–81.
Fevrier B, Vilette D, Archer F, Loew D, Faigle W, Vidal M, et al. Cells release prions in association with exosomes. Proc Natl Acad Sci U S A 2004; 101: 9683–8.
Grad LI, Pokrishevsky E, Silverman JM, Cashman NR . Exosome-dependent and independent mechanisms are involved in prion-like transmission of propagated Cu/Zn superoxide dismutase misfolding. Prion 2014; 8: 331–5.
Nonaka T, Masuda-Suzukake M, Arai T, Hasegawa Y, Akatsu H, Obi T, et al. Prion-like properties of pathological TDP-43 aggregates from diseased brains. Cell Rep 2013; 4: 124–34.
Zhang X, Yuan X, Shi H, Wu L, Qian H, Xu W . Exosomes in cancer: small particle, big player. J Hematol Oncol 2015; 8: 83.
Abd Elmageed ZY, Yang Y, Thomas R, Ranjan M, Mondal D, Moroz K, et al. Neoplastic reprogramming of patient-derived adipose stem cells by prostate cancer cell-associated exosomes. Stem Cells 2014; 32: 983–97.
Umezu T, Ohyashiki K, Kuroda M, Ohyashiki JH . Leukemia cell to endothelial cell communication via exosomal miRNAs. Oncogene 2013; 32: 2747–55.
Tadokoro H, Umezu T, Ohyashiki K, Hirano T, Ohyashiki JH . Exosomes derived from hypoxic leukemia cells enhance tube formation in endothelial cells. J Biol Chem 2013; 288: 34343–51.
Masyuk AI, Masyuk TV, Larusso NF . Exosomes in the pathogenesis, diagnostics and therapeutics of liver diseases. J Hepatol 2013; 59: 621–5.
Deng ZB, Liu Y, Liu C, Xiang X, Wang J, Cheng Z, et al. Immature myeloid cells induced by a high-fat diet contribute to liver inflammation. Hepatology 2009; 50: 1412–20.
Fleury A, Martinez MC, Le Lay S . Extracellular vesicles as therapeutic tools in cardiovascular diseases. Front Immunol 2014; 5: 370.
Corrado C, Raimondo S, Chiesi A, Ciccia F, De Leo G, Alessandro R . Exosomes as intercellular signaling organelles involved in health and disease: basic science and clinical applications. Int J Mol Sci 2013; 14: 5338–66.
Kosaka N, Iguchi H, Yoshioka Y, Takeshita F, Matsuki Y, Ochiya T . Secretory mechanisms and intercellular transfer of microRNAs in living cells. J Biol Chem 2010; 285: 17442–52.
Essandoh K, Yang L, Wang X, Huang W, Qin D, Hao J, et al. Blockade of exosome generation with GW4869 dampens the sepsis-induced inflammation and cardiac dysfunction. Biochim Biophys Acta 2015; 1852: 2362–71.
Al-Nedawi K, Meehan B, Kerbel RS, Allison AC, Rak J . Endothelial expression of autocrine VEGF upon the uptake of tumor-derived microvesicles containing oncogenic EGFR. Proc Natl Acad Sci U S A 2009; 106: 3794–9.
Lima LG, Chammas R, Monteiro RQ, Moreira ME, Barcinski MA . Tumor-derived microvesicles modulate the establishment of metastatic melanoma in a phosphatidylserine-dependent manner. Cancer Lett 2009; 283: 168–75.
Mulcahy LA, Pink RC, Carter DR . Routes and mechanisms of extracellular vesicle uptake. J Extracell Vesicles 2014; 3: 10.3402/jev.v3.24641.
Cai Z, Yang F, Yu L, Yu Z, Jiang L, Wang Q, et al. Activated T cell exosomes promote tumor invasion via Fas signaling pathway. J Immunol 2012; 188: 5954–61.
Lubas M, Christensen MS, Kristiansen MS, Domanski M, Falkenby LG, Lykke-Andersen S, et al. Interaction profiling identifies the human nuclear exosome targeting complex. Mol Cell 2011; 43: 624–37.
Lee RH, Pulin AA, Seo MJ, Kota DJ, Ylostalo J, Larson BL, et al. Intravenous hMSCs improve myocardial infarction in mice because cells embolized in lung are activated to secrete the anti-inflammatory protein TSG-6. Cell Stem Cell 2009; 5: 54–63.
Lai RC, Arslan F, Lee MM, Sze NS, Choo A, Chen TS, et al. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res 2010; 4: 214–22.
Ludwig AK, Giebel B . Exosomes: small vesicles participating in intercellular communication. Int J Biochem Cell Biol 2012; 44: 11–5.
Xin H, Li Y, Buller B, Katakowski M, Zhang Y, Wang X, et al. Exosome-mediated transfer of miR-133b from multipotent mesenchymal stromal cells to neural cells contributes to neurite outgrowth. Stem Cells 2012; 30: 1556–64.
Zhang B, Wang M, Gong A, Zhang X, Wu X, Zhu Y, et al. HucMSC-exosome mediated-wnt4 signaling is required for cutaneous wound healing. Stem Cells 2015; 33: 2158–68.
Ibrahim Ahmed GE, Cheng K, Marbán E . Exosomes as critical agents of cardiac regeneration triggered by cell therapy. Stem Cell Rep 2014; 2: 606–19.
Agarwal U, George A, Bhutani S, Ghosh-Choudhary S, Maxwell JT, Brown ME, et al. Experimental, systems, and computational approaches to understanding the microRNA-mediated reparative potential of cardiac progenitor cell-derived exosomes from pediatric patients. Circ Res 2017; 120: 701–12.
Hao ZC, Lu J, Wang SZ, Wu H, Zhang YT, Xu SG . Stem cell-derived exosomes: A promising strategy for fracture healing. Cell Prolif 2017; 50. doi:10.1111/cpr.12359.
Chen JY, An R, Liu ZJ, Wang JJ, Chen SZ, Hong MM, et al. Therapeutic effects of mesenchymal stem cell-derived microvesicles on pulmonary arterial hypertension in rats. Acta Pharmacol Sin 2014; 35: 1121–8.
Wu K, Yang Y, Zhong Y, Ammar HM, Zhang P, Guo R, et al. The effects of microvesicles on endothelial progenitor cells are compromised in type 2 diabetic patients via downregulation of the miR-126/VEGFR2 pathway. Am J Physiol Endocrinol Metab 2016; 310: E828–37.
Raemdonck K, Braeckmans K, Demeester J, De Smedt SC . Merging the best of both worlds: hybrid lipid-enveloped matrix nanocomposites in drug delivery. Chem Soc Rev 2014; 43: 444–72.
Li C, Zhang J, Zu YJ, Nie SF, Cao J, Wang Q, et al. Biocompatible and biodegradable nanoparticles for enhancement of anti-cancer activities of phytochemicals. Chin J Nat Med 2015; 13: 641–52.
Ha D, Yang N, Nadithe V . Exosomes as therapeutic drug carriers and delivery vehicles across biological membranes: current perspectives and future challenges. Acta Pharm Sin B 2016; 6: 287–96.
Kooijmans SA, Vader P, van Dommelen SM, van Solinge WW, Schiffelers RM . Exosome mimetics: a novel class of drug delivery systems. Int J Nanomedicine 2012; 7: 1525–41.
Sun D, Zhuang X, Xiang X, Liu Y, Zhang S, Liu C, et al. A novel nanoparticle drug delivery system: the anti-inflammatory activity of curcumin is enhanced when encapsulated in exosomes. Mol Ther 2010; 18: 1606–14.
Tian Y, Li S, Song J, Ji T, Zhu M, Anderson GJ, et al. A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy. Biomaterials 2014; 35: 2383–90.
Yang T, Martin P, Fogarty B, Brown A, Schurman K, Phipps R, et al. Exosome delivered anticancer drugs across the blood-brain barrier for brain cancer therapy in Danio rerio. Pharm Res 2015; 32: 2003–14.
Batrakova EV, Kim MS . Using exosomes, naturally-equipped nanocarriers, for drug delivery. J Control Release 2015; 219: 396–405.
Alvarez-Erviti L, Seow Y, Yin H, Betts C, Lakhal S, Wood MJ . Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol 2011; 29: 341–5.
Wahlgren J, De LKT, Brisslert M, Vaziri Sani F, Telemo E, Sunnerhagen P, et al. Plasma exosomes can deliver exogenous short interfering RNA to monocytes and lymphocytes. Nucleic Acids Res 2012; 40: e130.
Jiang XC, Gao JQ . Exosomes as novel bio-carriers for gene and drug delivery. Int J Pharm 2017; 521: 167–75.
Zhuang X, Xiang X, Grizzle W, Sun D, Zhang S, Axtell RC, et al. Treatment of brain inflammatory diseases by delivering exosome encapsulated anti-inflammatory drugs from the nasal region to the brain. Mol Ther 2011; 19: 1769–79.
Mignot G, Roux S, Thery C, Segura E, Zitvogel L . Prospects for exosomes in immunotherapy of cancer. J Cell Mol Med 2006; 10: 376–88.
Escudier B, Dorval T, Chaput N, Andre F, Caby MP, Novault S, et al. Vaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived-exosomes: results of thefirst phase I clinical trial. J Transl Med 2005; 3: 10.
Morse MA, Garst J, Osada T, Khan S, Hobeika A, Clay TM, et al. A phase I study of dexosome immunotherapy in patients with advanced non-small cell lung cancer. J Transl Med 2005; 3: 9.
El Kasmi KC, Holst J, Coffre M, Mielke L, de Pauw A, Lhocine N, et al. General nature of the STAT3-activated anti-inflammatory response. J Immunol 2006; 177: 7880–8.
H Rashed M, Bayraktar E, K Helal G, Abd-Ellah MF, Amero P, Chavez-Reyes A, et al. Exosomes: from garbage bins to promising therapeutic targets. Int J Mol Sci 2017; 18. pii: E538.
Vickers KC, Remaley AT . Lipid-based carriers of microRNAs and intercellular communication. Curr Opin Lipidol 2012; 23: 91–7.
Ohno S, Takanashi M, Sudo K, Ueda S, Ishikawa A, Matsuyama N, et al. Systemically injected exosomes targeted to EGFR deliver antitumor microRNA to breast cancer cells. Mol Ther 2013; 21: 185–91.
Akao Y, Iio A, Itoh T, Noguchi S, Itoh Y, Ohtsuki Y, et al. Microvesicle-mediated RNA molecule delivery system using monocytes/macrophages. Mol Ther 2011; 19: 395–9.
Haney MJ, Klyachko NL, Zhao Y, Gupta R, Plotnikova EG, He Z, et al. Exosomes as drug delivery vehicles for Parkinson's disease therapy. J Control Release 2015; 207: 18–30.
Wang J, Zheng Y, Zhao M . Exosome-based cancer therapy: implication for targeting cancer stem cells. Front Pharmacol 2016; 7: 533.
Meyer C, Losacco J, Stickney Z, Li L, Marriott G, Lu B . Pseudotyping exosomes for enhanced protein delivery in mammalian cells. Int J Nanomed 2017; 12: 3153–70.
Nordin JZ, Lee Y, Vader P, Mager I, Johansson HJ, Heusermann W, et al. Ultrafiltration with size-exclusion liquid chromatography for high yield isolation of extracellular vesicles preserving intact biophysical and functional properties. Nanomedicine 2015; 11: 879–83.
Lakhal S, Wood MJ . Exosome nanotechnology: an emerging paradigm shift in drug delivery: exploitation of exosome nanovesicles for systemic in vivo delivery of RNAi heralds new horizons for drug delivery across biological barriers. Bioessays 2011; 33: 737–41.
Li M, Zeringer E, Barta T, Schageman J, Cheng A, Vlassov AV . Analysis of the RNA content of the exosomes derived from blood serum and urine and its potential as biomarkers. Philos Trans R Soc Lond B Biol Sci 2014; 369: 20130502.
Batrakova EV, Kim MS . Development and regulation of exosome-based therapy products. Wiley Interdiscip Rev Nanomed Nanobiotechnol 2016; 8: 744–57.
Budnik V, Ruiz-Canada C, Wendler F . Extracellular vesicles round off communication in the nervous system. Nat Rev Neurosci 2016; 17: 160–72.
Elmore S . Apoptosis: a review of programmed cell death. Toxicol Pathol 2007; 35: 495–516.
Zhang J, Li S, Li L, Li M, Guo C, Yao J, et al. Exosome and exosomal microRNA: trafficking, sorting, and function. Genomics Proteomics Bioinformatics 2015; 13: 17–24.
Lee Y, El Andaloussi S, Wood MJA . Exosomes and microvesicles: extracellular vesicles for genetic information transfer and gene therapy. Hum Mol Genet 2012; 21: R125–34.
Chen B, Li Q, Zhao B, Wang Y . Stem cell-derived extracellular vesicles as a novel potential therapeutic tool for tissue repair. Stem Cells Transl Med 2017; 6: 1753–8.
Kajdos M, Janas L, Kolasa-Zwierzchowska D, Wilczynski JR, Stetkiewicz T . Microvesicles as a potential biomarker of neoplastic diseases and their role in development and progression of neoplasm. Prz Menopauzalny 2015; 14: 283–91.
Wlodkowic D, Telford W, Skommer J, Darzynkiewicz Z . Apoptosis and beyond: cytometry in studies of programmed cell death. Methods Cell Biol 2011; 103: 55–98.
Acknowledgements
This work was supported in part by an American Heart Association Grant-in-Aid 16GRNT30950010 and a National Institutes of Health COBRE grant P20GM104936 (to Johnson RAJASINGH).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Samanta, S., Rajasingh, S., Drosos, N. et al. Exosomes: new molecular targets of diseases. Acta Pharmacol Sin 39, 501–513 (2018). https://doi.org/10.1038/aps.2017.162
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/aps.2017.162
Keywords
This article is cited by
-
Exosome-mediated repair of spinal cord injury: a promising therapeutic strategy
Stem Cell Research & Therapy (2024)
-
Modification of immune cell-derived exosomes for enhanced cancer immunotherapy: current advances and therapeutic applications
Experimental & Molecular Medicine (2024)
-
Exploring the Role of Mesenchymal Stem Cell–Derived Exosomes in Diabetic and Chemotherapy-Induced Peripheral Neuropathy
Molecular Neurobiology (2024)
-
Exploring platelet-derived microvesicles in vascular regeneration: unraveling the intricate mechanisms and molecular mediators
Molecular Biology Reports (2024)
-
Neural stem cell-derived exosomes and regeneration: cell-free therapeutic strategies for traumatic brain injury
Stem Cell Research & Therapy (2023)
