
Abstract
The discovery of efficacious anti-ischemic drugs remains a challenge. Recently we have found that rosmarinic acid n-butyl ester (RABE), a derivative of rosmarinic acid, significantly protects SH-SY5Y cells against oxygen glucose deprivation (OGD)-induced cell death. In the present study we simultaneously investigated the effects of RABE on the two key players in the pathophysiology of cerebral ischemia, ischemic neuronal damage and microglial inflammation. Pretreatment with RABE (1, 10 μmol/L) dose-dependently attenuated OGD- or H2O2-induced reduction of the viability of SH-SY5Y neuroblastoma cells. RABE pretreatment concurrently reduced the apoptotic cell rate, down-regulated the expression of the pro-apoptotic proteins Bax and p53, and up-regulated the expression of the anti-apoptotic protein phosphorylated death-associated protein kinase (DAPK). Furthermore, pretreatment with RABE (3 μmol/L) markedly inhibited lipopolysaccharide (LPS)-induced increases in the release of TNF-α, IL-1β, NO and PGE2, and the expression levels of iNOS, and COX-2 in cultured rat microglial cells. In conclusion, these results reveal for the first time the potential anti-ischemic effects of RABE on neuronal and glial cells and elucidate the molecular mechanisms involved in its dual beneficial profiles in vitro. RABE may be a promising drug lead/candidate for the treatment of ischemic stroke.
Similar content being viewed by others

Introduction
Stroke is a multifactorial disease caused by a combination of environmental and genetic factors, and it is a major cause of serious long-term disability in adults and the second leading cause of death worldwide1,2. Approximately 87% of stroke patients are ischemic, and the remaining cases are hemorrhagic3. Ischemic strokes are caused by an interruption of the blood supply to the brain, while hemorrhagic strokes result from the rupture of a blood vessel or an abnormal vascular structure. Only recombinant tissue-plasminogen activator (rt-PA) for thrombolysis is approved by US Food and Drug Administration for use in the treatment of this devastating disease, but the use of this drug is restricted by a short treatment window4, a narrow eligible range5, and the risk of hemorrhage6. Effective therapeutic strategies remain a challenge despite the advances in our understanding of the pathophysiology of cerebral ischemia.
Ischemic stroke often occurs with complex pathophysiological events, and neuronal damage and microglial inflammation are two of the key players7,8,9,10. Neuronal death in the brain following ischemic injury is a dynamic and prolonged process that involves several apoptosis-related pathways, including the Fas signaling pathway11, notch pathway12, and p53 signaling pathway13. Death-associated protein kinase (DAPK) is a newly identified Ca2+/calmodulin-dependent kinase that plays an important role in the apoptosis process. A recent study demonstrated that DAPK was an irreplaceable regulator in ischemic neuronal death14,15. The DAPK-p53 interaction is a point of convergence during necrotic and apoptotic pathways, and this complex was considered a desirable target for the treatment of ischemic insults16. Cerebral ischemia also triggers a robust activation of microglial cells, which play pivotal roles in the acute and chronic phases of injury17,18. Microglia constitute approximately 12% of brain cells19, and these cells are responsible for inflammation, which is characterized into three stages after ischemic injury: acute, chronic and resolution20. Microglia largely produce pro-inflammatory cytokines, such as tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β), in the acute phase of brain inflammation in ischemic stroke21. Microglia-induced chronic inflammation leads to excessive release of nitric oxide (NO) by inducible nitric oxide synthase (iNOS)22,23,24 and prostaglandin E2 (PGE2) by cyclooxygenase-2 (COX-2)25. These inflammatory responses are substantial and detrimental factors in the pathogenesis of ischemic stroke26. There is a bidirectional interaction and communication between neurons and neighboring glial cells, and the amelioration of ischemia-associated neuronal damage and suppression of pro-inflammatory cytokines in activated microglia are generally accepted as important therapeutic regimens in the treatment of ischemic stroke.
Natural products are reliable resources in drug discovery and development processes27. For example, Butylphthalide (isolated from Apium graveolens)28 is used clinically for the treatment of stroke in China. in the current study, Rosmarinic acid (RA) and its synthetic derivatives (2–12) (Supplementary Chart S1) were investigated in oxygen-glucose deprivation (OGD) injury to identify the most effective agent as a new lead compound that is effective against ischemic stroke. Rosmarinic acid n-butyl ester (RABE) (5) was the most potent RA derivative, and it almost completely protected cells against OGD-induced reduction in the survival of SH-SY5Y cells (Supplementary Figure S1). We further demonstrated the dual protective effects of RABE against ischemia-associated neuronal damage and inflammatory responses of microglial cells and investigated the potential molecular mechanisms underlying this beneficial profile.
Materials and methods
RABE preparation
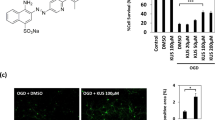
RABE (Figure 1) is a derivative of rosmarinic acid. The RABE employed in the present study was provided by Prof Wei-min ZHAO of the Shanghai Institute of Materia Medica, Chinese Academy of Sciences. The synthetic procedures of RABE and purity reports are provided in the Supplementary Information. RABE purity was above 95% using high-performance liquid chromatography (HPLC) analysis [HPLC: column, Athena C18-WP, 21 mm×50 mm; solvent system, acetonitrile-0.2‰ HCOOH with gradient elution; flow rate, 0.3 mL/min; UV detection, 327 nm; Shimadzu LC-MS-2020, Shimadzu Corporation, Kyoto, Japan]. RABE was dissolved in dimethylsulfoxide (DMSO) to 10 mmol/L as a stock solution and stored at -20 °C. The stock solution was diluted with culture medium before use. Cell cultures were pre-incubated with different concentrations of RABE for 2 h prior to exposure to various insults.

Chemical structure of RABE.
Cell culture
Human neuroblastoma SH-SY5Y cells were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA) and cultured in a 1:1 mixture of Eagle's minimum essential medium and Ham's F12 medium (MEM/F12; Gibco, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS; Gibco, Grand Island, NY, USA), 100 U/mL penicillin and 100 μg/mL streptomycin. The cells were seeded into 96-well plates or 6-well plates (Corning, NY, USA) at a density of 2×105 cells/mL and maintained at 37 °C in 95% air/5% CO2 in a humidified incubator. The culture media were replaced with fresh media 24 h after seeding for H2O2 stimulation, or with the DMEM medium for OGD insult.
Primary microglia were prepared from the cortex of Sprague Dawley (SD) rats at postnatal d 1 using the “shaking-off” method as described previously30. Briefly, mechanically dissociated cortical cells were seeded in 75-cm2 T-flasks (1 brain/flask) containing DMEM/F12 medium (Gibco, Grand Island, NY, USA) supplemented with 10% FBS, 100 U/mL penicillin and 100 μg/mL streptomycin at 37 °C in 95% air/5% CO2 in a humidified incubator with the medium replaced every three days. Microglia floated off and separated from mixed glial cultures on d 14 in vitro, and these cells were seeded into 96-well plates or 6-well plates.
Hydrogen peroxide (H2O2), OGD, or lipopolysaccharide (LPS) insults
H2O2 was freshly prepared from an 8.8-mol/L stock solution prior to each experiment, and the final concentration in the culture medium was 100 μmol/L for the entire study. The cells used for OGD exposure were washed with OGD medium (DMEM medium without glucose) and placed in OGD medium in a hypoxia chamber (85% N2/10% H2/5% CO2 at 37 °C) for 1 h. The cells were placed in normal DMEM medium (supplied with 1 g/L glucose and 10% FBS) after OGD insults and returned to a standard incubator (95% air/5% CO2 at 37 °C) for another 24 h. The culture medium of the control group was changed to normal DMEM medium at the same time, and these cells were incubated for another 24 h. Microglia were incubated with 100 ng/mL LPS for 24 h to induce inflammatory responses. Assays were performed at different times after exposure, as specified in the following methods.
Cell viability assay
Cell viability was evaluated using morphological observation under a microscope (Nikon TE2000, Melville, NY, USA) and the 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sigma, St Louis, MO, USA) assay. When obtained by the live cell, MTT is transformed to formazan, and the amount of formazan directly correlates with the number of metabolically active cells31. MTT was added to the culture media at a final concentration of 0.5 mg/mL after different treatments and incubated at 37 °C for 3 h. The formed formazan was dissolved by the addition of 100 μL DMSO to each well. Cell plates were shaken for 5 min, and the absorbance of each well was recorded on a DTX 800 Multimode Detector (Beckman Coulter, Fullerton, CA, USA) at 490 nm.
Lactate dehydrogenase (LDH) assay
LDH is a stable cytosolic enzyme that is released upon cell lysis, and it is used to measure cytotoxicity. The supernatant medium was collected 24 h after H2O2 or OGD exposure and centrifuged at 300×g for 10 min to pellet debris. The supernatant medium was used for LDH assay, which was performed according to the LDH kit manufacturer's instructions (Nanjing Jiancheng, Nanjing, China). The absorbance was measured at a wavelength of 450 nm using a DTX 800 Multimode Detector (Beckman Coulter, Fullerton, CA, USA).
Flow cytometry analysis
Annexin V-FITC and propidium iodide (PI) staining (BD Biosciences, San Jose, CA, USA) was used to identify apoptotic cells. Annexin V-FITC staining precedes the loss of membrane integrity that accompanies the latest stages of cell death resulting from apoptotic or necrotic processes, while PI identifies early apoptotic cells. SH-SY5Y cells were collected via centrifugation at 300×g for 10 min. The cells were washed twice with cold PBS and resuspended in 100 μL (1×105 cells) of binding buffer. Annexin V-FITC (5 μL) and PI (5 μL) were added to the binding buffer and incubated for 15 min at room temperature in the dark. Another 400 μL of binding buffer was added to the samples, and cells were analyzed using flow cytometry (BD Biosciences, San Jose, CA, USA) within 1 h.
Nuclear fraction extraction
The primary contribution of p53 to apoptosis in response to ischemic injury is dependent on transcriptional activity. In order to test the effect of RABE on the transcriptional activation of p53, nuclear fraction extraction was performed to assess changes in protein from the cytoplasm to nucleus. Cells were harvested using trypsin-EDTA and centrifuged at 500×g for 5 min. Nuclear extracts were obtained using NE-PER Reagents (Thermo Scientific, USA) for Western blotting.
Western blot analysis
Protein levels were determined using immunoblotting. Cells were homogenized in RIPA buffer (50 mmol/L Tris-base, 150 mmol/L NaCl, 0.5% sodium deoxycholate, 1% NP-40, 0.1% SDS, pH 7.5) with protease inhibitors and centrifuged at 12 000×g for 10 min at 4 °C. The protein concentrations in supernatants were determined using a bicinchoninic acid (BCA) assay kit (Pierce, Rockford, IL, USA). After addition of loading buffer, protein samples were electrophoresed on 8%–12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene difluoride (PVDF) membranes (0.45 μm; Millipore, Billerica, MA, USA). The membranes were blocked with 5% skimmed milk in T-TBS (0.05% (v/v) Tween-20, 50 mmol/L Tris-base, 150 mmol/L, pH 7.5) at room temperature for 1 h and probed with primary anti-Bax (1:30 000; Abcam, Cambridge, UK), anti-p53 (1:2000; Cell Signaling, Beverly, MA, USA), anti-p-DAPK, anti-DAPK (1:1000; Sigma, St Louis, MO, USA), anti-iNOS (1:1000; BD Biosciences, San Jose, CA, USA), anti-GAPDH (1:10 000; Kangcheng, Shanghai, China), and anti-lamin B (1:5000; Cell Signaling, Beverly, MA, USA), or anti-α-tubulin (1:5000; Santa Cruz, CA, USA) antibodies at 4 °C overnight, followed by incubation with a horseradish peroxidase (HRP)-conjugated secondary antibody (1:5000; Kangcheng, Shanghai, China) for 1 h at room temperature. Membranes were washed and developed using an enhanced chemiluminescence (ECL) reagent (Millipore, Billerica, MA, USA) according to the manufacturer's instructions and visualized using autoradiography film. Band intensity was quantified using Image J software.
NO release assay
The LPS-induced lease of NO in the cultured supernatants was determined by measuring Nitrite (NO2–) concentrations, a major stable product of NO. The nitrite concentrations were measured using the Griess reaction. Each sample (50 μL) was reacted with an equal volume of Griess reagent in 96-well plates for 15 min at room temperature in the dark. The absorbance of the mixture at 540 nm was determined using a DTX 800 Multimode Detector (Beckman Coulter, Fullerton, CA, USA). Nitrite concentrations were calculated from a sodium nitrite standard curve.
Measurement of inflammatory cytokines
Culture media were collected 24 h after LPS treatment. The samples were centrifuged at 1000×g for 10 min. The supernatants were collected and stored at -80 °C until analysis. TNF-α, IL-1β and PGE2 protein levels were quantified using ELISA kits (Invitrogen, Carlsbad, CA, USA) according to manufacturer's instructions. Absorbance at 450 nm was recorded using a DTX 800 Multimode Detector (Beckman Coulter, Fullerton, CA, USA). Cytokine concentrations were normalized to protein content, which was measured using a BCA Protein Assay Kit.
Statistical analysis
All data were collected from at least three independent experiments. The data are expressed as the mean±SEM. Statistical analyses of differences between groups were analyzed using one-way ANOVA followed by Tukey's test for multiple comparisons or Student's t-test for single comparisons. Statistical significance was established at P value <0.05.
Results
Neuroprotective effects of RABE on SH-SY5Y cells
Two harmful ischemic stimuli, OGD and H2O2, were used to stimulate SH-SY5Y neuronal cells to investigate the neuroprotective effects of RABE. OGD or H2O2 stimulation caused a significant reduction in cell viability, as observed by morphological changes and MTT and LDH assays. SH-SY5Y cells exposed to OGD for 1 h followed by reoxygenation for 24 h or H2O2 for 24 h resulted in cell morphological changes, such as neurite disappearance, body shrinkage and reduced cell numbers (Figure 2A and 2D). In contrast, cell morphological changes induced by OGD or H2O2 were remarkably attenuated by pre-treatment with 1 or 10 μmol/L of RABE (Figure 2A and 2D). Ischemia-induced cell damage was also demonstrated by a significant MTT reduction (48.86%±3.17%, P<0.01; 39.58%±3.93%, P<0.01, respectively) (Figure 2B and 2E) and an excessive release of LDH (207.33%±11.35%, P<0.01; 139.00%±4.78%, P<0.01, respectively) (Figure 2C and 2F) compared to the control group. However, RABE pretreatment (1 and 10 μmol/L) exhibited a dose-response attenuation of OGD or H2O2-induced cell viability reduction with maximum protection at 10 μmol/L (restored viability to 96.26%±4.69%, P<0.01 vs OGD group; 86.70%±0.51%, P<0.01 vs H2O2 group) (Figure 2B and 2E). RABE pretreatment of SH-SY5Y cells reduced LDH levels only at 10 μmol/L (140.33%±20.61%, P<0.05 vs OGD group) (Figure 2C) when exposed to OGD insult. RABE at 1 and 10 μmol/L decreased LDH to 121.00%±3.61% (P<0.05 vs H2O2 group) and 117.17%±3.70% (P<0.01 vs H2O2 group) (Figure 2F), respectively, in the H2O2 model. RABE at 1 and 10 μmol/L exerted no obvious toxic effects on SH-SY5Y cells with 24 h of incubation (Supplementary Figure S2).

Effects of RABE on SH-SY5Y cells exposed to OGD or H2O2 injury. SH-SY5Y cells were pre-treated with 1 or 10 μmol/L of RABE followed by exposing to OGD for 1 h followed by reoxygenation for 24 h or H2O2 for 24 h. Phase-contrast micrographs (×100) of SH-SY5Y cells after different RABE treatment with OGD (A) or H2O2 (D) insult. Cell viability was measured by MTT assay after RABE pretreatment with OGD (B) or H2O2 (E) injury. LDH in the culture medium was measured after RABE treatment with exposure to OGD (C) or H2O2 (F) injury. The data were presented as mean±SEM from three independent experiments. **P<0.01 vs control. #P<0.05, ##P<0.01 vs OGD or H2O2 group.
The anti-apoptotic effects of RABE on SH-SY5Y cells
Flow cytometry using annexin V-FITC and PI staining was performed to examine whether the neuroprotective effects of RABE in SH-SY5Y cells corresponded to reduced apoptosis. The distribution of SH-SY5Y cells to the lower left quadrant suggested that these cells were viable and without measurable apoptosis. OGD and H2O2 insults induced obvious increases in cell numbers in the upper right quadrants, which indicated that these SH-SY5Y cells underwent late apoptosis (Figure 3A and 3C, respectively). The percentage of SH-SY5Y cells at early and late stages of apoptosis increased from 8.19%±2.40% to 23.90%±3.45% (P<0.01) (Figure 3B), in the OGD group compared to the control group. The percentage of apoptotic cells increased significantly from 9.09%±0.84% to 24.98%±2.75% after exposure to H2O2 (Figure 3D, P<0.01 vs control). Pretreatment with RABE at 10 μmol/L suppressed the apoptotic ratio to 10.46%±1.93% (Figure 3B, P<0.05 vs OGD group), and 1 μmol/L exhibited a trend of attenuation of the apoptotic rate, but it was not statistically significant. RABE decreased the percentage of apoptotic rate to 16.36%±1.65% (P<0.05 vs H2O2 group) and 12.51%±2.59% (P<0.05 vs H2O2 group) at 1 and 10 μmol/L, respectively (Figure 3D).

Effect of RABE on apoptosis in SH-SY5Y cells induced by OGD or H2O2. SH-SY5Y cells were pre-treated with the indicated concentration of RABE for 2 h before stimulating with OGD or H2O2 for 24 h. Representative plots of flow cytometric analysis for OGD and H2O2-treated cells respectively (A, C), which shows of the percentage of apoptotic cells: the sum of the upper right and lower right quadrants. The percentage of apoptosis was showed by the histogram (B, D), which was from results of three independent experiments. **P<0.01 vs control. #P<0.05 vs OGD or H2O2 group.
The DAPK-p53 signaling pathway was involved in RABE neuroprotection of SH-SY5Y cells
The influence of RABE on DAPK, p53, and Bax expression were examined to elucidate the potential molecular mechanism involved in the anti-apoptotic effects of RABE. OGD and H2O2 injury markedly induced Bax expression (62.75%±13.58%, P<0.05 compared to the OGD group; 61.33%±6.06%, P<0.01 compared to the H2O2 group) (Figure 4A and 4D), p53 nuclear translocation (20.60%±10.23%, P<0.01 compared to the OGD group; 17.33%±3.71%, P<0.01 compared to the H2O2 group) (Figure 4B and 4E), and DAPK dephosphorylation (33.40%±14.22%, P<0.01; 28.60%±10.13%, P<0.01 compared to the control group) (Figure 4C and 4F). RABE at 1 and 10 μmol/L remarkably down-regulated OGD- or H2O2-induced Bax (64.00%±7.02%, P<0.01, 52.75%±7.48%, P<0.01 compared to the OGD group; 54.33%±12.55%, P<0.05, 46.00%±17.10%, P<0.05 compared to the H2O2 group) and p53 over-expression (69.20%±11.16%, P<0.05, 36.20%±11.84%, P<0.01 compared to the OGD group; 32.33%±14.77%, P<0.05, 12.33%±7.36%, P<0.01 compared to the H2O2 group) (Figure 4A, 4B, 4D and 4E) and significantly reversed the dephosphorylation of DAPK following OGD or H2O2 exposure only at 10 μmol/L (74.80%±17.10%, P<0.05 compared to the OGD group; 95.40%±21.02%, P<0.05 compared to the H2O2 group) (Figure 4C and 4F).

Effect of RABE on Bax, p53 and phosphorylated DAPK expression in SH-SY5Y cells after OGD or H2O2 injury. SH-SY5Y cells were pre-incubated with RABE and exposed to OGD or H2O2 for 24 h. The expression of Bax (A, D), p53 (B, E) and p-DAPK/DAPK (C, F) were determined by Western blot analysis. The histogram of the protein expression percentage was the results of three independent experiments. **P<0.01 vs control. #P<0.05, ##P<0.01 vs OGD or H2O2 group.
RABE decreased the production of acute pro-inflammatory cytokines in LPS-stimulated primary microglia
LPS-stimulated primary microglia were used to investigate the effects of RABE on inflammatory responses. ELISAs demonstrated that LPS (100 ng/mL) exposure for 24 h resulted in increased TNF-α (549.00±44.23 pg/mL, P<0.01 vs control) (Figure 5A) and IL-1β (239.45±9.95 pg/mL, P<0.01 vs control) (Figure 5B) release in the supernatants of primary microglia cultures. Pre-incubation with 3 μmol/L RABE significantly suppressed the increase in TNF-α and IL-1β production to 357.74±52.46 pg/mL (P<0.05 vs LPS group) (Figure 5A) and 99.89±37.36 pg/mL (P<0.05 vs LPS group) (Figure 5B), respectively. No significant changes in cell viability were observed in primary microglia after incubation with 3 μmol/L of RABE or 100 ng/mL of LPS for 24 h (Supplementary Figure S3).

Effect of RABE on the production of TNF-α and IL-1β in culture media of LPS-stimulated primary microglia. After pre-treatment with 3 μmol/L RABE, primary microglia were exposed to LPS for 24 h. The levels of TNF-α (A) and IL-1β (B) were assessed by ELISA assay. Values were expressed as mean±SEM (n=3). **P<0.01 vs control. #P<0.05 vs LPS group.
RABE inhibited of nitrite production and iNOS expression in LPS-stimulated primary microglia
LPS stimulation for 24 h resulted in increased nitrite release in the supernatants of primary microglia cultures (9.81±1.07 μmol/L, P<0.01) compared to the control (2.46±0.48 μmol/L) (Figure 6A). Treatment with 3 μmol/L of RABE reduced nitrite production to 4.13±0.92 μmol/L (P<0.05 vs LPS group). We investigated the expression of iNOS, which is responsible for the release of NO, to delineate the mechanism of RABE-mediated inhibition of LPS-induced NO release. LPS stimulation markedly up-regulated iNOS expression (control was 5.00%±1.53% of LPS group, P<0.01), and RABE treatment at 3 μmol/L significantly decreased iNOS levels (20.50%±12.50%, P<0.01 vs LPS group) (Figure 6B).

Effect of RABE on LPS-stimulated nitrite production and iNOS expression in LPS-induced primary microglia. Cells were pretreated with RABE for 2 h before stimulation of LPS for another 24 h. The release of nitrite was analyzed by a Griess reaction in culture media (A). The expression of iNOS was analyzed by Western blot (B). Values were expressed as mean±SEM of three independent experiments. **P<0.01 vs control. #P<0.05, ##P<0.01 vs LPS group.
RABE reduced PGE2 levels and COX-2 expression in LPS-stimulated primary microglia
An ELISA was used to investigate the effect of RABE on PGE2 production in LPS-induced microglia. Exposure of primary microglia to LPS increased the secretion of PGE2 (1260.47±151.80 pg/mL, P<0.01) compared to the control (215.00±36.08 pg/mL) (Figure 7A). However, 3 μmol/L of RABE treatment significantly suppressed LPS-induced over-expression of PGE2 (667.83±39.47 pg/mL, P<0.01 vs LPS group) (Figure 7A). We measured changes in COX-2 expression, which is an upstream enzyme for PGE2 production, to further investigate the mechanism underlying the effect of RABE on PGE240. LPS induced a robust increase in COX-2 levels (control is 26.33%±6.12% of LPS group, P<0.01), and 3 μmol/L of RABE treatment reduced the increased expression of COX-2 (85.00%±1.73%, P<0.01 vs LPS group) in primary microglia (Figure 7B). We also observed that the cell viability of neurons was significantly decreased after exposure to LPS-exposed culture medium of microglia (primary microglia-condition medium, CM) for 24 h. In contrast, culture medium from LPS-exposed microglia with RABE treatment did not cause notable damage to the survival of primary neurons (Supplementary Figure S4).

Effect of RABE on the release of PGE2 and the expression of COX-2 in LPS-induced primary microglia. Primary microglia were treated with RABE for 2 h, followed by the stimulation of LPS for 24 h. After incubation, the culture media were collected and analyzed by ELISA assay for PGE2 production (A). Cell lysates were subjected to Western blot for COX-2 measurement (B). Results were expressed as mean±SEM (n=3). **P<0.01 vs control. ##P<0.01 vs LPS group.
Discussion
Accumulating evidence indicates that therapies modulating single target generally do not achieve satisfactory efficacy in the treatment of complex disease, including ischemic stroke. Alternatively, substantial effort has been exerted to discover potent small molecules that target multiple key cellular signaling pathways to produce superior therapeutic effects32. The present study identified a rosmarinic acid derivative, RABE, that exhibited dual antagonistic effects against neuronal damage and glial inflammation via concurrent regulation of apoptotic pathways and reduction of inflammatory factors. To our knowledge, this study is the first report of a rosmarinic acid-skeleton small molecule that directly ameliorated neuronal and glial dysfunction in separate cellular systems.
An OGD-stimulated SH-SY5Y neuronal model was used to mimic the lack of energy supply in neurons after ischemic insult in the current study33,34, and an H2O2-stimulated SH-SY5Y neuronal model was used to mimic the influence of ischemia-caused oxidative insult to neurons35. RABE exhibited marked protection against OGD- and H2O2-stimulated neuronal damage, as indicated by MTT and LDH measurements (Figure 1). These protective effects of RABE occurred concurrently with its amelioration of apoptotic damage induced by the same stimuli (Figure 2). Neuronal death is generally considered the result of necrosis or apoptosis. The LDH assay is used to assess the loss of cell membrane integrity associated with necrosis36, and MTT measurement reflects neuronal viability, which is affected by apoptosis or necrosis36,37,38. Therefore, the beneficial roles of RABE on the two ischemic-neuronal models suggest that RABE treatment may simultaneously suppress ischemic-associated necrotic and apoptotic neuronal deaths.
The potential molecular mechanisms of the anti-apoptotic activity of RABE were further investigated by assessing apoptosis-associated pathway. Apoptosis is an important mechanism of cell death after ischemia insults. DAPK is an important regulator of neuronal death, including apoptosis14,15. DAPK is phosphorylated in the resting state, but OGD induces DAPK dephosphorylation39. A previous study demonstrated that activated DAPK catalyzed direct binding with p53, which may produce a transcription-dependent pathway of apoptosis16. The primary roles of p53 in cell apoptosis involve the transcriptional induction of pro-apoptotic genes, such as Bax40. Our study demonstrated that RABE inhibited Bax over-expression, p53 translocation and DAPK dephosphorylation.
Later events of ischemia-reperfusion, such as the microglia-induced inflammatory responses, are widely propagated to exacerbate neuronal death after ischemic stroke attacks41. Microglia are the resident immune cells that mediate inflammation via the over-production of various pro-inflammatory cytokines, such as TNF-α and IL-1β, in the central nervous system (CNS) during the acute stage21. A chronically activated state of microglia releases toxic amounts of NO via iNOS22,23,24 and PGE2 via COX-225, which have a deleterious effect on post-ischemic inflammation. LPS is the most common toxic agent used to induce inflammation in primary microglia42, and this agent was used to investigate the effects of RABE on inflammatory responses. We found that RABE inhibited TNF-α and IL-1β release from LPS-stimulated primary microglia. RABE also suppressed LPS-induced NO and PGE2 over-production in primary microglia, which may be mediated by iNOS and COX-2 inhibition. The anti-inflammatory effect of RABE was further demonstrated by the protection of primary neurons against toxicity of a culture medium derived from LPS-exposed microglia.
In summary, the primary finding of the present study is the novel identification of an active rosmarinic acid analogue, RABE, which exhibited dual protective effects on ischemia-induced neuronal damage and microglial inflammation (Figure 8). We demonstrated the potent protective effects of RABE on neuronal cells against two different ischemic insults, OGD or H2O2 injury. These beneficial profiles of RABE were closely associated with its anti-apoptotic effect, which may be partially attributed to the DAPK-p53 signaling pathway. We also observed a significant suppressive effect of RABE on microglial inflammation via the simultaneous inhibition of acute-phase inflammatory responses, such as TNF-α and IL-1β over-production, and chronic-phase inflammatory responses, including NO and PGE2 release, which may occur via regulation of the upstream enzymes, iNOS and COX-2, respectively. Taken together, our study suggests that the dual anti-ischemic effects of RABE on neuronal and glial damage and the novel potential mechanisms provide important support for the development of drug leads/candidates against ischemic stroke. However, solid evidence of the efficacy of the post-treatment effects of RABE in primary cultured neurons, microglia and ischemia-reperfusion animal models, and the precise mechanisms of the beneficial aspects of this compound, are necessary to make a firm recommendation for future research.

Schematic diagram of the beneficial effects and potential mechanisms of RABE against ischemic insults. RABE dose-dependently decreased OGD or H2O2-induced ischemic neuronal damages, which may be mediated by downregulation of pro-apoptotic proteins Bax and p53, as well as upregulation of anti-apoptotic protein phosphorylated death-associated protein kinase (DAPK). Meanwhile, RABE also inhibited LPS-induced acute and chronic inflammation. These dual beneficial effects of RABE may synergistically interfere the ischemic damages.
References
Leifer D . Stroke. In: Binder MD, Hirokawa N, Windhorst U, editors. Encyclopedia of Neuroscience. Academic Press: Oxford; 2009. p 573–8.
Feigin VL, Forouzanfar MH, Krishnamurthi R, Mensah GA, Connor M, Bennett DA, et al. Global and regional burden of stroke during 1990-2010: findings from the Global Burden of Disease Study 2010. Lancet 2014; 383: 245–54.
Go AS, Mozaffarian D, Roger VL, Benjamin EJ, Berry JD, Blaha MJ, et al. Executive summary: heart disease and stroke statistics--2014 update: a report from the American Heart Association. Circulation 2014; 129: 399–410.
Hacke W, Kaste M, Bluhmki E, Brozman M, Dávalos A, Guidetti D, et al. Thrombolysis with alteplase 3 to 4.5 hours after acute ischemic stroke. N Engl J Med 2008; 359: 17–29.
Del Zoppo GJ, Saver JL, Jauch EC, Adams HP Jr, American Heart Association Stroke Council. Expansion of the time window for treatment of acute ischemic stroke with intravenous tissue plasminogen activator: a science advisory from the American Heart Association/American Stroke Association. Stroke 2009; 40: 2945–8.
Tanne D, Kasner SE, Demchuk AM, Koren-Morag N, Hanson S, Grond M, et al. Markers of increased risk of intracerebral hemorrhage after intravenous recombinant tissue plasminogen activator therapy for acute ischemic stroke in clinical practice: the Multicenter rt-PA Stroke Survey. Circulation 2002; 105: 1679–85.
Dirnagl U, Iadecola C, Moskowitz MA . Pathobiology of ischaemic stroke: an integrated view. Trends Neurosci 1999; 22: 391–7.
Brouns R, De Deyn PP . The complexity of neurobiological processes in acute ischemic stroke. Clin Neurol Neurosurg 2009; 111: 483–95.
Prestigiacomo CJ, Kim SC, Connolly ES Jr, Liao H, Yan SF, Pinsky DJ . CD18-mediated neutrophil recruitment contributes to the pathogenesis of reperfused but not nonreperfused stroke. Stroke 1999; 30: 1110–7.
Drake C, Boutin H, Jones MS, Denes A, McColl BW, Selvarajah JR, et al. Brain inflammation is induced by co-morbidities and risk factors for stroke. Brain Behav Immun 2011; 25: 1113–22.
Wajat H . The Fas signaling pathway: more than a paradigm. Science 2002; 296: 1635–6.
Artavanis-Tsakonas S, Rand MD, Lake RJ . Notch signaling: cell fate control and signal integration in development. Science 1999; 284: 770–6.
Moll UM, Wolff S, Speidel D, Deppert W . Transcription-independent pro-apoptotic functions of p53. Curr Opin Cell Biol 2005; 17: 631–6.
He C, Stroink AR, Wang CX . The role of DAPK-BimEL pathway in neuronal death induced by oxygen-glucose deprivation. Neuroscience 2014; 258: 254–62.
Fujita Y, Yamashita T . Role of DAPK in neuronal cell death. Apoptosis 2014; 19: 339–45.
Pei L, Shang Y, Jin H, Wang S, Wei N, Yan H, et al. DAPK1-p53 interaction converges necrotic and apoptotic pathways of ischemic neuronal death. J Neurosci 2014; 34: 6546–56.
Amantea D, Nappi G, Bernardi G, Bagetta G, Corasaniti MT . Post-ischemic brain damage: pathophysiology and role of inflammatory mediators. FEBS J 2009; 276: 13–26.
Wang Q, Tang XN, Yenari MA . The inflammatory response in stroke. J Neuroimmunol 2007; 184: 53–68.
Block ML, Zecca L, Hong JS . Microglia-mediated neurotoxicity: uncovering the molecular mechanisms. Nat Rev Neurosci 2007; 8: 57–69.
Brown GC, Neher J . Inflammatory neurodegeneration and mechanisms of microglial killing of neurons. J Mol Neurobiol 2010; 41: 242–7.
Lambertsen KL, Biber K, Finsen B . Inflammatory cytokines in experimental and human stroke. J Cereb Blood Flow Metab 2012; 32: 1677–98.
Forster C, Clark HB, Ross ME, Iadecola C . Inducible nitric oxide synthase expression in human cerebral infarcts. Acta Neuropathol 1999; 97: 215–20.
Moro MA, Cárdenas A, Hurtado O, Leza JC, Lizasoain I . Role of nitric oxide after brain ischaemia. Cell Calcium 2004; 36: 265–75.
Murphy S, Gibson CL . Nitric oxide, ischaemia and brain inflammation. Biochem Soc Trans 2007; 5: 1133–7.
Kawano T, Anrather J, Zhou P, Park L, Wang G, Frys KA, et al. Prostaglandin E2 EP1 receptors: downstream effectors of COX-2 neurotoxicity. Nat Med 2006; 12: 225–9.
Kawabori M, Yenari MA . Inflammatory responses in brain ischemia. Curr Med Chem 2015; 22: 1258–77.
Newman DJ, Cragg GM . Natural products as sources of new drugs from 1981 to 2014. J Nat Prod 2016; 79: 629–61.
Zhang L, Yu WH, Wang YX, Wang C, Zhao F, Qi W, et al. DL-3-n-Butylphthalide, an anti-oxidant agent, prevents neurological deficits and cerebral injury following stroke per functional analysis, magnetic resonance imaging and histological assessment. Curr Neurovasc Res 2012; 9: 167–75.
Xue LX, Zhang T, Zhao YW, Geng Z, Chen JJ, Chen H . Efficacy and safety comparison of DL-3-n-butylphthalide and Cerebrolysin: Effects on neurological and behavioral outcomes in acute ischemic stroke. Exp Ther Med 2016; 11: 2015–20.
Gordon R, Hogan CE, Neal ML, Anantharam V, Kanthasamy AG, Kanthasamy AJ . A simple magnetic separation method for high-yield isolation of pure primary microglia. Neurosci Meth 2011; 194: 287–96.
Hansen MB, Nielsen SF, Berg KJ . Re-examination and further development of a precise and rapid dye method for measuring cell growth/cell kill. Immunol Methods 1989; 119: 203–10.
Chen HS, Qi SH, Shen JG . One-compound-multi-target: combination prospect of natural compounds with thrombolytic therapy in acute ischemic stroke. Curr Neuropharmacol 2017; 15: 134–56.
Liu Y, Lu Z, Cui M, Yang Q, Tang Y, Dong Q . Tissue kallikrein protects SH-SY5Y neuronal cells against oxygen and glucose deprivation-induced injury through bradykinin B2 receptor-dependent regulation of autophagy induction. J Neurochem 2016; 139: 208–20.
Jiang CS, Guo XJ, Gong JX, Zhu TT, Zhang HY, Guo YW . Synthesis and biological evaluation of 21-arylidenepregnenolone derivatives as neuroprotective agents. Bioorg Med Chem Lett 2012; 22: 2226–9.
Wang XJ, Wang LY, Fu Y, Wu J, Tang XC, Zhao WM, et al. Promising effects on ameliorating mitochondrial function and enhancing Akt signaling in SH-SY5Y cells by (M)-bicelaphanol A, a novel dimeric podocarpane type trinorditerpene isolated from Celastrus orbiculatus. Phytomedicine 2013; 20: 1064–70.
Hankø E, Hansen TW, Almaas R, Lindstad J, Rootwelt T . Bilirubin induces apoptosis and necrosis in human NT2-N neurons. Pediatr Res 2005; 57: 179–84.
Tripathy D, Grammas P . Acetaminophen inhibits neuronal inflammation and protects neurons from oxidative stress. J Neuroinflammation 2009; 6: 10.
Aldasoro M, Guerra-Ojeda S, Aguirre-Rueda D, Mauricio MD, Vila JM, Marchio P, et al. Effects of ranolazine on astrocytes and neurons in primary culture. PLoS One 2016; 11: e0150619.
Shohat G, Spivak-Kroizman T, Cohen O, Bialik S, Shani G, Berrisi H, et al. The pro-apoptotic function of death-associated protein kinase is controlled by a unique inhibitory autophosphorylation-based mechanism. Biol Chem 2001; 276: 47460–7.
Amaral JD, Xavier JM, Steer CJ, Rodrigues CM . Targeting the p53 pathway of apoptosis. Curr Pharm Des 2010; 16: 2493–503.
Chavez JC, Hurko O, Barone FC, Feuerstein GZ . Pharmacologic interventions for stroke: looking beyond the thrombolysis time window into the penumbra with biomarkers, not a stopwatch. Stroke 2009; 40: e558–63.
Weinstein JR, Swarts S, Bishop C, Hanisch UK, Möller T . Lipopolysaccharide is a frequent and significant contaminant in microglia-activating factors. Glia 2008; 56: 16–26.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (No 81522045 and 81473113) and the State Key Laboratory of Drug Research, Shanghai Institute of Materia Medica, Chinese Academy of Sciences, Shanghai, China (SIMM1601KF-03).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Supplementary information is available at website of Acta Pharmacologica Sinica.
Supplementary information
Supplementary Chart S1
Structures of rosmarinic acid (1) and its analogues. (PDF 332 kb)
Supplementary Figure S1
Effect of rosmarinic acid (1) and its analogues (2-12) on SH-SY5Y cells subjected to OGD. (PDF 273 kb)
Supplementary Figure S2
RABE had no effect on cell viability in SH-SY5Y cells. (PDF 273 kb)
Supplementary Figure S3
Effect of RABE or LPS on cell viability in primary microglia. (PDF 264 kb)
Supplementary Figure S4
RABE protected primary neurons from primary microglia-condition medium (CM) injury. (PDF 389 kb)
Rights and permissions
About this article

Cite this article
Wu, L., Wang, Hm., Li, Jl. et al. Dual anti-ischemic effects of rosmarinic acid n-butyl ester via alleviation of DAPK-p53-mediated neuronal damage and microglial inflammation. Acta Pharmacol Sin 38, 459–468 (2017). https://doi.org/10.1038/aps.2016.156
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/aps.2016.156
Keywords
This article is cited by
-
Study on the Role and Mechanism of Exosomes Derived from Dental Pulp Stem Cells in Promoting Regeneration of Myelin Sheath in Rats with Sciatic Nerve Injury
Molecular Neurobiology (2024)
-
Neuroprotective Effects of Sodium Butyrate through Suppressing Neuroinflammation and Modulating Antioxidant Enzymes
Neurochemical Research (2021)
