
Abstract
2,3,5,4′-Tetrahydroxystilbene-2-O-β-D-glucoside (TSG) is a water-soluble active component extracted from Polygonum multiflorum Thunb. A number of studies demonstrate that TSG exerts cardioprotective effects. Since endoplasmic reticulum (ER) stress plays a key role in myocardial ischemia/reperfusion (MI/R)-induced cell apoptosis, we sought to determine whether modulation of the ER stress during MI/R injury was involved in the cardioprotective action of TSG. Male mice were treated with TSG (60 mg·kg−1·d−1, ig) for 2 weeks and then were subjected to MI/R surgery. Pre-administration of TSG significantly improved post-operative cardiac function, and suppressed MI/R-induced myocardial apoptosis, evidenced by the reduction in the myocardial apoptotic index, serum levels of LDH and CK after 6 h of reperfusion. TSG (0.1–1000 μmol/L) did not affect the viability of cultured H9c2 cardiomyoblasts in vitro, but pretreatment with TSG dose-dependently decreased simulated ischemia/reperfusion (SIR)-induced cell apoptosis. Furthermore, both in vivo and in vitro studies revealed that TSG treatment activated the Notch1/Hes1 signaling pathway and suppressed ER stress, as evidenced by increasing Notch1, Notch1 intracellular domain (NICD), Hes1, and Bcl-2 expression levels and by decreasing p-PERK/PERK ratio, p-eIF2α/eIF2α ratio, and ATF4, CHOP, Bax, and caspase-3 expression levels. Moreover, the protective effects conferred by TSG on SIR-treated H9c2 cardiomyoblasts were abolished by co-administration of DAPT (the Notch1 signaling inhibitor). In summary, TSG ameliorates MI/R injury in vivo and in vitro by activating the Notch1/Hes1 signaling pathway and attenuating ER stress-induced apoptosis.
Similar content being viewed by others

Introduction
Cardiovascular diseases are the leading contributor to mortality rates worldwide. Myocardial ischemia is one of the main consequences of cardiovascular diseases1. Because of the lack of effective therapies, MI/R injury remains a major medical problem today2. Timely reperfusion is essential for recovery of the ischemic myocardium; however, reperfusion itself results in major cardiac damage3,4. The underlying mechanism regulating myocardial injury induced by MI/R remains to be fully elucidated.
The endoplasmic reticulum (ER) is a type of organelle in the cells of eukaryotic organisms whose membranes are continuous with the outer membrane of the nuclear envelope. ER participates in the biosynthesis and folding of proteins5. Many physiological and pathological stimulations, such as ischemia, hypoxia, and poisons, induce the overexpression of ER molecular chaperones and an unfolded protein response (UPR) to increase normal protein folding and decrease the abnormal protein folding through a process referred to as ER-associated degradation (ERAD)6,7. UPR signaling promotes survival and adaptation of ER stress. However, when UPR activation is excessive or prolonged, the final cellular outcome is pathologic apoptotic cell death8. MI/R injury is known to result in the accumulation of unfolded proteins in the ER, which causes a severe UPR. Persistent UPR eventually leads to cellular apoptosis. Therefore, reducing excessive UPR, also referred to as ER stress, is important in ameliorating MI/R injury9.
2,3,5,4'-Tetrahydroxystilbene-2-O-β-D-Glucoside (TSG, molecular formula: C20H22O9, molecular weight: 406, Figure 1), a water-soluble active component extracted from Polygonum multiflorumThunb, exhibits a wide spectrum of biological functions, such as anti-inflammatory, anti-oxidant, anti-aging, lipid lowering, neuroprotection, and cardioprotection effects10,11,12,13. Recently, TSG has attracted much attention as a therapeutic agent against a number of heart diseases, including atherosclerosis, myocardial fibrosis, cardiac hypertrophy and vascular senescence14,15,16,17. Importantly, it has been demonstrated that many organs suffering from ischemic injury can be ameliorated by TSG18. Based on its cardioprotective properties, TSG has elicited substantial interest as a potential agent to attenuate MI/R injury19. Nevertheless, the specific mechanism of TSG-mediated cardioprotection remains largely unknown.

Chemical structure of TSG. Molecular weight: 406. Molecular formula: C20H22O9.
The Notch signaling pathway regulates multiple cellular processes, including cell fate determination, differentiation, development, proliferation, apoptosis and regeneration20,21. In mammals, 4 Notch receptors (Notch1–4) and five Notch ligands (Delta-like1/3/4, and Jagged1/2) have been identified21. The activation of Notch receptor occurs via binding to Notch ligands. The target genes of Notch signaling include basic helix-loop-helix transcription factors Hairy and enhancer of split 1 (Hes1) and Hairy-related transcription (HRT) factor family members, which are found in both developing and adult hearts22. In the heart, Notch signaling not only regulates embryonic cardiac development and differentiation23, but also stimulates proliferation of immature cardiomyocytes24 and promotes quiescent cardiomyocytes to reenter the cell cycle25. Many studies have shown that the Notch1, NICD and the downstream target protein Hes1, which decrease in cardiomyocytes during cardiac postnatal development, are restored and activated in the adult injured myocardium26. However, whether TSG protects cardiomyocytes against MI/R-induced ER stress and myocardial apoptosis through a Notch1-dependent mechanism is not yet known.
In this study, TSG was used as an adjuvant to attenuate the MI/R-induced ER stress and myocardial apoptosis. In addition, the involvement of the Notch signaling pathway in the protective actions of TSG was evaluated.
Materials and methods
Animals
Male C57Bl/6 mice (6–8 weeks of age, weighing 20–25 g) were obtained from the animal center of the Fourth Military Medical University. All procedures were approved by the Ethics Committee of the Fourth Military Medical University and performed according to the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (National Institutes of Health Publication No 85-23, revised in 1996).
Materials
TSG (over 98% pure) was obtained from the Department of Chemistry, the Fourth Military Medical University. DAPT were purchased from Abcam (Cambridge, MA, USA). Dulbecco's modified Eagle's medium (DMEM) and fetal bovine serum (FBS) were purchased from Gibco (Waltham, MA, USA). Penicillin and streptomycin were purchased from Beijing Solarbio Science & Technology Co, Ltd (Beijing, China). Cell Counting kit-8 (CCK-8) was purchased from Dojindo Laboratories Co, Ltd (Shanghai, China). A terminal deoxynucleotidyl-transferase-mediated dUTP nick-end labeling (TUNEL) assay kit and protease inhibitor cocktail were obtained from Roche Molecular Biochemicals (Mannheim, Germany). Lactate dehydrogenase (LDH) and creatine kinase (CK) assay kits were obtained from Jiancheng Bioengineering Institute (Nanjing, China). A bicinchoninic acid (BCA) protein quantification kit was purchased from Merck Millipore Technology (Darmstadt, Germany). Primary antibodies against Notch1, NICD, Hes1, and CHOP were obtained from Abcam (Cambridge, MA, USA); p-PERK, PERK, p-eIF2α, eIF2α, and β-actin were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA); and ATF4, caspase-3, Bax, and Bcl-2 were obtained from Cell Signaling Technology (Danvers, MA, USA). Secondary antibodies were purchased from the Zhongshan Company (Beijing, China).
Experimental protocol
Step 1 was designed to evaluate the effect of TSG on MI/R injured hearts. C57Bl/6 mice were orally gavaged with TSG at a dose of 60 mg/kg body weight per day for 2 weeks. The TSG dose was selected based on previous studies16,27,28. Two weeks after the treatment, MI/R surgery was performed as previously described29. Briefly, C57Bl/6 mice were anesthetized with 1% pentobarbital sodium. The myocardial ischemia operation was performed by exteriorizing the heart through a left thoracic incision, placing a 6–0 silk suture around the left anterior descending coronary artery and tying a slipknot. After 30 min of ischemia, the slipknot was released and the myocardium was reperfused for 4 h (for analysis of protein expression), 6 h (for quantification of myocardial apoptosis), and 72 h (for cardiac function determination). Mice in the sham group underwent the same operation procedures except that the suture that passed under the left coronary artery was left untied.
Step 2 was designed to investigate whether TSG pretreatment alleviated SIR injury in H9c2 cardiomyoblasts, which improved the cellular viability and reduced apoptosis. The present study first evaluated the effect of TSG on normal H9c2 cardiomyoblasts. The TSG stock solution was prepared in heated PBS solution and diluted with DMEM immediately prior to use. The cells were randomly divided into control and TSG (0.1, 1, 10, 100, or 1000 μmol/L) groups. Cell viability was then detected using the CCK-8 assay. Then, the H9c2 cardiomyoblasts were randomly divided into control, SIR, and TSG (1, 10, or 100 μmol/L)+SIR groups. TSG (1, 10, or 100 μmol/L)+SIR groups were separately incubated with TSG for 8 h prior to SIR treatment in vitro. The effects of 0.1 and 1000 μmol/L TSG on H9c2 cardiomyoblasts were also explored. The CCK-8 assay demonstrated that 0.1 μmol/L TSG had almost no effect on improving the cell viability (compared with the SIR group, P>0.05), whereas the effect of 1000 μmol/L TSG on improving the cell viability was no better than the effect of 100 μmol/L TSG. As a result, 1, 10, and 100 μmol/L TSG were selected for this study. Then, the H9c2 cardiomyoblasts were treated with SIR in the absence or presence of TSG or DAPT (5 μmol/L) treatment. The DAPT dose was selected based on previous studies30,31. The cells were harvested after SIR treatment for further analysis.
Echocardiography
Two-dimensional and M-mode echocardiographic measurements were performed with a VEVO 770 high-resolution in vivo imaging system (VisualSonics, Toronto, ON, Canada) 72 h after MI/R operation, as previously described32.
Determination of myocardial apoptosis
Myocardial apoptosis was analyzed by a TUNEL (terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling) assay using an in situ cell death detection kit, as described previously33. The apoptosis index was expressed as the number of apoptotic myocytes/the total number of myocytes counted ×100%.
Determination of serum CK and LDH
Blood samples (1 mL) were collected after 6 h of reperfusion. Serum CK and LDH levels were spectrophotometrically measured (Beckman DU 640, Fullerton, CA, USA) according to the manufacturer's instructions34.
Cell culture
H9c2 embryonic rat myocardium-derived cells, a well-characterized and widely used cell line to study myocardial cell ischemia, were cultured in DMEM supplemented with 10% fetal bovine serum (FBS) and 1% antibiotic-antimycotic. The H9c2 cardiomyoblasts were maintained at 37 °C with 5% CO2. For all treatment groups, the H9c2 cardiomyoblasts (100 000 cells/mL) were pre-incubated with or without different TSG concentrations (1, 10, and 100 μmol/L) or DAPT (5 μmol/L) for 8 h prior to initiating SIR treatment.
Simulated ischemia/reperfusion treatment
SIR treatment was performed using physiological concentrations of potassium, hydrogen, and lactate. The procedure was performed as previously described35. Briefly, the H9c2 cardiomyoblasts were exposed to an ischemic buffer containing (in mmol/L) 137 NaCl, 12 KCl, 0.49 MgCl2, 0.9 CaCl2, and 4 HEPES. This buffer was also supplemented with (in mmol/L) 10 deoxyglucose, 0.75 sodium dithionate, and 20 lactate. The buffer pH was 6.5, and the cells were incubated for 50 min in a humidified cell culture incubator (21% O2, 5% CO2, 37 °C). Reperfusion was initiated by returning the cells to normal culture medium for 4 h in a humidified cell culture incubator (21% O2, 5% CO2, 37 °C).
Cell viability assay
After H9c2 cardiomyoblasts were seeded in 96-well culture plates and received different treatments, 10 μL of CCK-8 solution was added to each well at a 1/10 dilution. After 2 h of incubation, the absorbance was measured using a microtiter plate reader (SpectraMax 190, Molecular Device, USA) at a wavelength of 450 nm. The cell viability was calculated by dividing the optical density of samples with the optical density of the control group. In addition, the cell morphology was observed with an inverted/phase contrast microscope, and images were obtained (Olympus BX61, Japan).
Determination of cellular apoptosis
After H9c2 cardiomyoblasts were fixed in paraformaldehyde (4%) for 24 h, the cellular apoptosis was also analyzed by performing a TUNEL assay using the in situ cell death detection kit according to the manufacturer's instructions31. The apoptotic index was expressed as the number of positively stained apoptotic cells/the total number of cells counted ×100%.
Western blot analysis
Myocardial tissue and cultured cells were harvested and lysed after the indicated treatments. The lysates were clarified by centrifugation at 4 °C for 10 min at 12 000 r/min. After quantitation of the protein concentration, 30 μg of total protein was separated by SDS-PAGE and then transferred to a poly vinylidene difluoride membrane (Millipore, USA). The membrane was blocked with 5% free-fat milk in TBST for 2 h at room temperature and then incubated with primary antibodies specific to Notch1, NICD, Hes1, caspase-3, Bcl-2, Bax, ATF4, CHOP, p-PERK, PERK, p-eIF2α, eIF2α, and β-actin (1:500) overnight at 4 °C. After washing three times with TBST, the membrane was incubated with secondary antibodies for 2 h at room temperature, and they were then washed as above. The positive protein bands were developed using a chemiluminescent system, and the bands were scanned and quantified by densitometric analysis using an image analyzer Quantity One System (Bio-Rad, Richmond, CA, USA).
Statistical analysis
Data are expressed as the mean±SEM. Differences were compared by ANOVA followed by Bonferroni correction for a post hoc t test, where appropriate. P<0.05 was considered to be statistically significant. All statistical tests were performed with the GraphPad Prism software version 5.0 (GraphPad Software, Inc, San Diego, CA, USA).
Results
The effect of TSG treatment on MI/R injury
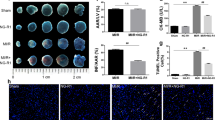
To investigate whether TSG has cardioprotective effects against MI/R in mice, the present study first examined the left ventricular function after 72 h of reperfusion. Echocardiography showed an increased left ventricular ejection fraction (LVEF) and left ventricular fractional shortening (LVFS) in the TSG-treated group compared with the MI/R group (Figure 2, P<0.05). To examine whether TSG decreased myocardial apoptosis, we further measured the myocardial apoptotic index, serum LDH and CK levels after 6 h of reperfusion. Compared with the MI/R group, TSG treatment attenuated cardiac apoptosis by decreasing the apoptotic index (Figure 3, P<0.05). In addition, the serum CK and LDH levels were also significantly reduced after 6 h of reperfusion in the TSG treatment group (Figure 3, P<0.05, compared with the MI/R group).

The effect of TSG on cardiac function in MI/R injured hearts. (A) Representative M-mode images by echocardiography. Cardiac function was assessed by echocardiography 72 h after MI/R operation. (B) Left ventricular ejection fraction (LVEF). (C) Left ventricular fractional shortening (LVFS). The results are expressed as the mean±SEM. n=8/group. *P<0.05 vs the sham group. #P<0.05 vs the MI/R group. MI/R, myocardial ischemia/reperfusion (30 min/72 h).

The effect of TSG treatment on apoptotic index, lactate dehydrogenase (LDH) release, and serum creatine kinase (CK) release in MI/R injured hearts. (A) Left: representative photomicrographs of in situ detection of apoptotic myocytes by TUNEL staining. Green fluorescence shows TUNEL-positive nuclei and blue fluorescence shows nuclei of total cardiomyocytes; original magnification, ×400. Right: percentage of TUNEL-positive nuclei; (B) Serum lactate dehydrogenase (LDH) level; and (C) Serum creatine kinase (CK) level. The results are expressed as the mean±SEM. n=8/group. *P<0.05 vs the sham group. #P<0.05 vs the MI/R group. MI/R, myocardial ischemia/reperfusion (30 min/6 h).
The effect of TSG treatment on the Notch1/Hes1 signaling pathway and ER stress in MI/R hearts
We further measured the levels of Notch1/Hes1 signaling and the expression of ER stress and apoptotic related proteins in MI/R injured hearts. As depicted in Figure 4, Western blotting analysis indicated that MI/R significantly increased the ratios of p-PERK/PERK and p-eIF2α/eIF2α as well as the expression of ATF4, CHOP, caspase-3 and Bax compared with the Sham group; conversely, MI/R produced a significant decrease in the expression of Bcl-2 (P<0.05, compared with the Sham group). Treatment with TSG produced a significant increase in the Notch1, NICD, Hes1 and Bcl-2 expression levels (P<0.05, compared with the MI/R group). Additionally, TSG treatment also down-regulated the ratios of p-PERK/PERK and p-eIF2α/eIF2α as well as the ATF4, CHOP, caspase-3 and Bax expression levels (P<0.05, compared with the MI/R group).

The effect of TSG treatment on the Notch1/Hes1 signaling levels and the expression of ER stress and apoptotic related proteins in MI/R injured hearts. (A) Representative blots; (B) Notch1 expression; (C) NICD expression; (D) Hes1 expression; (E) p-PERK/PERK ratio; (F) p-eIF2α/eIF2α ratio; (G) ATF4 expression; (H) CHOP expression; (I) Bcl-2 expression; (J) Bax expression; and (K) Caspase-3 expression. The results are expressed as the mean±SEM. n=8/group. *P<0.05 vs the sham group. #P<0.05 vs the MI/R group. MI/R, myocardial ischemia/reperfusion (30 min/4 h).
The effect of TSG treatment on cell viability and apoptosis in SIR-treated H9c2 cardiomyoblasts
To investigate whether TSG was able to protect against cardiac injury in vitro, the present study first determined the effect of TSG on the survival of SIR-treated H9c2 cardiomyoblasts. The H9c2 cardiomyoblasts were exposed to TSG (0.1, 1, 10, 100, or 1000 μmol/L) for 8 h. Then, cell viability was determined by CCK-8. As shown in Figure 5A, no significant cell viability change was found compared with the Control group (P>0.05). Next, the cells were subjected to SIR treatment in the absence or presence of TSG (0.1, 1, 10, 100, or 1000 μmol/L) treatment. SIR treatment significantly decreased the cell viability (Figure 5B, P<0.05, compared with the Control group), while TSG (1, 10, or 100 μmol/L) treatment significantly increased the cell viability in a dose-dependent manner (Figures 5B and 5C, P<0.05, compared with the SIR group). However, 0.1 μmol/L TSG exhibited almost no effect on improving the cell viability (compared with the SIR group, P>0.05); the effect of 1000 μmol/L TSG on improving the cell viability was not better than the effect of 100 μmol/L TSG.

Effect of TSG treatment on cell viability and apoptosis in SIR-treated H9c2 cardiomyoblasts. In cultured H9c2 cardiomyoblasts subjected to SIR treatment, TSG significantly reduced the cellular apoptosis. H9c2 cardiomyoblasts were exposed to TSG treatment for 8 h at the concentrations of 1, 10, and 100 μmol/L, separately. Then, cell viability was measured by CCK-8. (A–B) Cell viability was determined by CCK-8 and was calculated by dividing the optical density of samples with the optical density of the control group; (C) Cell morphology was observed under an inverted phase contrast microscope and images were obtained; and (D) Top: representative photomicrographs of TUNEL staining. The green florescence shows the TUNEL-positive nuclei and the blue florescence shows the nuclei of all cells; original magnification, ×200. Bottom: percentage of TUNEL-positive nuclei. The results are expressed as the mean±SEM of 8 separate experiments with triplicate samples for each experimental condition within each experiment. *P<0.05 vs the control group. #P<0.05 vs the SIR group. nsP>0.05 vs the SIR group.
Treatment with TSG prevented the cell apoptosis induced by SIR treatment. To examine whether TSG has any effect on SIR-induced apoptosis in H9c2 cardiomyoblasts, the present study performed a TUNEL assay. The nuclei of normal cells were stained by propidium iodide and those of apoptotic cells were stained green. The number of TUNEL-positive cells was manually counted in five randomly selected fields, with the apoptotic index expressed as a percentage of the total counted cells (Figure 5D). TUNEL staining showed that SIR treatment increased the apoptotic index (P<0.05, compared with the Control group, Figure 5D). Compared with the SIR group, TSG treatment attenuated H9c2 cardiomyoblast apoptosis by decreasing apoptotic index in a dose-dependent manner (P<0.05, Figure 5D).
The effect of TSG treatment on caspase-3, Bax, Bcl-2, and ER stress-related protein expression in SIR-treated H9c2 cardiomyoblasts
To investigate whether the protective effect of TSG is associated with the apoptosis induced by ER stress signaling during SIR injury on H9c2 cardiomyoblasts, the protein levels of pro-apoptotic caspase-3, Bax, and anti-apoptotic Bcl-2, and the ER stress response proteins p-PERK, PERK, p-eIF2α, eIF2α, ATF4, and CHOP were measured using Western blot analysis. CHOP is a critical pro-apoptotic factor in ER stress-associated apoptosis. As shown in Figure 6, the present study found that the protein expression of CHOP in the SIR group was significantly up-regulated compared with that in the Control group (P<0.05). However, TSG attenuated the SIR-induced up-regulation in the expression of CHOP compared with the SIR group (P<0.05). In addition, as depicted in Figure 6, Western blot analysis indicated that SIR significantly increased the ratios of p-PERK/PERK and p-eIF2α/eIF2α as well as the expression levels of ATF4, caspase-3, and Bax (P<0.05, compared with the Control group); conversely, SIR produced a significant decrease in the Bcl-2 expression (P<0.05, compared with the Control group). However, TSG treatment down-regulated the ratios of p-PERK/PERK and p-eIF2α/eIF2α as well as the expression levels of ATF4, caspase-3, and Bax (P<0.05, compared with the SIR group). Additionally, treatment with TSG also produced a significant increase in Bcl-2 expression (P<0.05, compared with the SIR group).

In cultured H9c2 cardiomyoblasts subjected to SIR treatment, TSG significantly attenuated the PERK/eIF2α/ATF4-mediated ER stress and down-regulated the cell apoptosis. (A) Representative blots; (B) p-PERK/PERK ratio; (C) p-eIF2α/eIF2α ratio; (D) ATF4 expression; (E) CHOP expression; (F) Bcl-2 expression; (G) Bax expression; and (H) Caspase-3 expression. The results are expressed as the mean±SEM. n=8/group. *P<0.05 vs the control group. #P<0.05 vs the SIR group.
The effect of TSG treatment on Notch1, NICD and Hes1 expression in SIR-treated H9c2 cardiomyoblasts
The findings of our previous experiments indicated that TSG attenuated ER stress and protected H9c2 cardiomyoblasts from SIR-induced cell death. To explore the role of the Notch1/Hes1 signaling pathway in the protective action of TSG, we further measured the expression levels of Notch1, NICD, and Hes1. As depicted in Figure 7, moderate levels of Notch1, NICD and Hes1 were detected in the Control group, and these protein levels were slightly increased following SIR treatment. Notably, a significant increase in the Notch1, NICD, and Hes1 protein levels was observed in the TSG+SIR group (P<0.05).

The effect of TSG on Notch1, NICD, and Hes1 expression in SIR-treated H9c2 cardiomyoblasts. (A) Representative blots; (B) Notch1 expression; (C) NICD expression; and (D) Hes1 expression. The results are expressed as the mean±SEM. n=8/group. *P<0.05 vs the SIR group.
The effects of TSG and DAPT co-treatment on cell viability and apoptosis in SIR-treated H9c2 cardiomyoblasts
To further investigate the potential role of the Notch1/Hes1 signaling pathway in the protective effect of TSG, we determined whether DAPT treatment affected the cell viability (which was increased by TSG) in SIR-treated H9c2 cardiomyoblasts. The cells were subjected to SIR treatment in the absence or presence of TSG (100 μmol/L). SIR treatment significantly decreased cell viability, as in our previous experiments (Figure 5, P<0.05, compared with the Control group), while TSG pretreatment for 8 h resulted in a significant increase in the cell viability (Figure 8, P<0.05, compared with the SIR group). However, co-treatment with DAPT blocked the protective effect of TSG (Figure 8, P<0.05, compared with the SIR+TSG group). As observed under a microscope, TSG treatment significantly attenuated SIR-induced cell shrinkage and improved cell attachment. However, this protective effect was partially blunted by co-treatment with DAPT (Figure 8A). As shown in Figure 8C, TSG treatment also reduced the apoptotic index, while DAPT co-treatment significantly blocked this protective effect (P<0.05, compared with the SIR+TSG group).

In cultured H9c2 cardiomyoblasts subjected to SIR treatment, TSG significantly reduced the cellular apoptosis, whereas DAPT markedly blocked these effects. (A) Cell morphology was observed under an inverted phase contrast microscope and images were obtained; (B) The cell viability was evaluated using CCK-8; and (C) Left: representative photomicrographs of TUNEL staining, the green florescence shows the TUNEL-positive nuclei and the blue florescence shows the nuclei of all cells; original magnification, ×200. Right: percentage of TUNEL-positive nuclei. The results are expressed as the mean±SEM. n=8/group. *P<0.05 vs the SIR group. #P<0.05 vs the SIR+TSG group.
The effects of TSG and DAPT co-treatment on Notch1, NICD, and Hes1 expression in SIR-treated H9c2 cardiomyoblasts
To verify that TSG activated Notch1/Hes1 signaling in H9c2 cardiomyoblasts, we further measured the effects of TSG and DAPT on the expression levels of Notch1, NICD, and Hes1 in cultured H9c2 cardiomyoblasts that were exposed to SIR. Consistently, significant up-regulation of Notch1, NICD and Hes1 expression levels was found in the TSG treatment group (Figure 9, P<0.05, compared with the SIR group), while co-treatment with DAPT blocked Notch1/Hes1 activation, which was induced by TSG (Figure 9, P<0.05, compared with the SIR+TSG group).

In cultured H9c2 cardiomyoblasts subjected to SIR treatment, TSG effectively up-regulated the Notch1/Hes1 signaling, whereas DAPT blocked these effects. (A) Representative blots; (B) Notch1 expression; (C) NICD expression; and (D) Hes1 expression. The results are expressed as the mean±SEM. n=8/group. *P<0.05 vs the SIR group. #P<0.05 vs the SIR+TSG group. $P<0.05 vs the SIR+TSG+DAPT group.
The effects of TSG and DAPT co-treatment on caspase-3, Bax, Bcl-2, and ER stress-related protein expression in SIR-treated H9c2 cardiomyoblasts
In cultured H9c2 cardiomyoblasts, SIR injury significantly enhanced PERK/eIF2α/ATF4/CHOP-mediated ER stress, which was markedly suppressed by TSG treatment. However, compared with the SIR+TSG group, DAPT co-treatment significantly increased the expression levels of p-PERK/PERK, p-eIF2α/eIF2α, ATF4, and CHOP (all P<0.05, Figure 10), indicating that DAPT inhibited the ameliorative effect of TSG on SIR-induced ER stress. In addition, DAPT also blocked the anti-apoptotic effects of TSG by increasing the expression of caspase-3 and Bax and decreasing the Bcl-2 expression (Figure 10, P<0.05, compared with the SIR+TSG group).

In cultured H9c2 cardiomyoblasts subjected to SIR treatment, TSG significantly attenuated the PERK/eIF2α/ATF4-mediated ER stress and down-regulated the cell apoptosis, whereas DAPT abolished these effects. (A) Representative blots; (B) p-PERK/PERK ratio; (C) p-eIF2α/eIF2α ratio; (D) ATF4 expression; (E) CHOP expression; (F) Bcl-2 expression; (G) Bax expression; and (H) Caspase-3 expression. The results are expressed as the mean±SEM. n=8/group. *P<0.05 vs the SIR group. #P<0.05 vs the SIR+TSG group. $P<0.05 vs the SIR+TSG+DAPT group.
Discussion
In this study, we utilized in vivo and in vitro models to investigate the protective effect of TSG against MI/R-induced cardiac damage. We found that TSG treatment conferred cardioprotective effects, as evidenced by improved post-MI/R cardiac functional recovery, attenuated ER stress and reduced myocardial apoptosis. Importantly, Notch1/Hes1 signaling was proven to play a key role in this process. This study could help further elucidate the pharmacology of TSG and highlighted a novel strategy against ischemic heart disease.
The ER is responsible for synthesizing, modifying, folding and trafficking secretory proteins as well as regulating intracellular Ca2+ homoeostasis. It is highly sensitive to the deprivation of cellular energy and alterations of the redox state or Ca2+ concentration. These changes lead to the accumulation of unfolded proteins, a condition called ER stress36. The ER stress pathway was first identified as a cellular response that is induced by the accumulation of unfolded proteins in the ER to preserve the organelle's function. Previous studies have demonstrated that ER stress is one of the main events in the pathogenesis of MI/R injury37,38,39,40. Activation of the UPR in mammalians is mediated by three distinct ER stress sensors, namely, ER kinase (PERK), transcription factor ATF6, and endoribonuclease IRE-1. In these three pathways, IRE1 and ATF6 rectify and re-establish ER homeostasis, while PERK eliminates misfolded proteins to elicit pro-apoptotic effects in case ER homeostasis is not reinstated41,42. Therefore, the most immediate response to ER stress is provoked by PERK43. In addition, it has been well established that the C/EBP homologous protein (CHOP) pathway is the main pathway through which apoptosis is induced during ER stress44,45. MI/R injury activates ER stress, causing apoptosis by promoting the expression of CHOP9. CHOP is downstream of the PERK-eIF2α-ATF4 pathway in UPR46. The activated, phosphorylated form of PERK (p-PERK) widely suppresses the synthesis of functional proteins in cells by phosphorylating its target protein, the α-subunit of eukaryotic initiation factor 2 (eIF2α), which then enhances translation of ATF4, induces the expression of CHOP, and actively promotes cell apoptosis. A previous study demonstrated that inhibition of PERK/eIF2α-mediated ER stress reduced the MI/R injury in rat hearts47. In this study, we found that TSG decreased the phosphorylation levels of PERK and eIF2α as well as the expression of ATF4 and CHOP, reducing the ER stress caused by the MI/R injury in vivo and in vitro. To the best of our knowledge, this was to determine the role of ER stress in TSG-mediated cardioprotection against MI/R injury.
Notch1/Hes1 signaling has indispensable functional activities to control tissue formation in cardiogenesis48. The binding of the Notch1 ligand to its receptor triggers the γ-secretase-mediated proteolytic cleavage of NICD, which then translocates into the nucleus, up-regulating its downstream target gene Hes149,50. Notch signaling is believed to play a key role in maintaining functional homeostasis in adult hearts, as has been shown in studies related to heart models, such as myocardial infarction, cardiac hypertrophy and failure26,51. Interestingly, as the myocardium matures to the adult hormonal and contractile state, endogenous Notch signaling levels decreases steadily in heart tissue24,26. However, accumulating evidence has indicated that re-activation of Notch signaling could constitute an adaptive response to the increased survival rate or regenerate tissues following pathological stress. For example, activation of endogenous Notch following myocardial infarction in the adult can promote anti-apoptosis, increase the survival rate and improve the cardiac performance48. When Notch signaling was blocked in newborn mice by the administration of DAPT, the mortality rate tended to increase. After DAPT cessation, cardiac function was largely restored52. Furthermore, a previous study showed that activation of Notch1 signaling in H9c2 cardiomyoblasts contributed to the cardioprotection provided by ischemic preconditioning and postconditioning53. Pei et al demonstrated that Notch1 signaling protected against reperfusion injury in murine ischemic myocardium54. These studies indicated the key role of Notch signaling in maintaining functional homeostasis in adult hearts. In this study, we found that the expression levels of Notch1, NICD, and Hes1 were slightly up-regulated by MI/R injury and SIR treatment, suggesting endogenous activation of cardiac Notch1 signal during MI/R injury and SIR treatment, whereas a significant increase in these proteins levels was observed in the TSG-treated group, indicating the critical role of Notch1/Hes1 signaling in the protective action of TSG.
DAPT, a potent γ-secretase inhibitor, has been widely used to specifically block the Notch1/Hes1 signaling by preventing the intracellular domain of Notch (NICD) from being released and then translocated into the nucleus. In the present study, co-treatment with DAPT significantly attenuated TSG's protective effect against SIR injury on H9c2 cardiomyoblasts, indicating that Notch1/Hes1 signaling played a pivotal role under these conditions. Interestingly, a recent study found that γ-secretase inhibitor triggered apoptosis of primary chronic lymphocytic leukemia cells by inhibiting proteasome activity and enhancing as well as amplifying ER stress signaling by blocking Notch activation55. This finding suggested that inhibition of the Notch1/Hes1 signaling could enhance ER stress signaling, which somewhat coincided with our experimental results. In this study, we found that Notch1/Hes1 signaling activation by TSG reduced the ER stress level. However, blocking this pathway by DAPT attenuated this effect. Additionally, Ohta et al found that ER stress could enhance γ-secretase activity and increase the expression of Notch1 target genes56. Another study showed that Musashi genes regulated insulin expression, apoptosis and proliferation in response to ER stress via Hes157. These findings suggested that ER stress might also regulate Notch1/Hes1 signaling. Although further study is needed to investigate the detailed mechanisms, we demonstrated that TSG reduced myocardial ER stress by activating Notch1/Hes1 signaling.
Interestingly, Notch1 signaling exhibited different functions under different pathological conditions. Our previous study demonstrated that Notch1 signaling is a crucial mediator of endothelial inflammatory damage31 and TSG exerts a solid protective effect against endothelial inflammation by inhibiting the Notch signaling pathway58. However, Notch1 signaling plays a different role in cardiomyocytes. In the present study, we found that TSG activates Notch1/Hes1 signaling, thus reducing MI/R injury. The diverse role of Notch1 signaling may be due to the different models and pathological conditions in these two studies. Further study is definitely needed to investigate the effect of TSG on Notch signaling in other tissues and conditions.
In summary, our findings showed that TSG reduces myocardial ER stress via activating Notch1/Hes1 signaling, thus ameliorating MI/R injury. These findings indicate that TSG may be a promising candidate for treating ischemic heart diseases.
Author contribution
The work presented here was performed in collaboration among all authors. Si-wang WANG and Wei-xun DUAN defined the research theme and critically revised the manuscript. Meng ZHANG, Li-ming YU, and Hang ZHAO designed the methods and experiments, performed the laboratory experiments and wrote the paper. Xuan-xuan ZHOU, Qian YANG, Fan SONG, Meng-en ZHAI, Bu-ying LI, Li YAN, Bin ZHANG, and Zhen-xiao JIN collected and analyzed the data and interpreted the results. All authors have read and approved the final manuscript.
References
Di Diego JM, Antzelevitch C . Acute myocardial ischemia: cellular mechanisms underlying ST segment elevation. J Electrocardiol 2014; 47: 486–90.
Penna C, Perrelli MG, Tullio F, Angotti C, Camporeale A, Poli V, et al. Diazoxide postconditioning induces mitochondrial protein S-nitrosylation and a redox-sensitive mitochondrial phosphorylation/translocation of RISK elements: no role for SAFE. Basic Res Cardiol 2013; 108: 371.
Logue SE, Gustafsson AB, Samali A, Gottlieb RA . Ischemia/reperfusion injury at the intersection with cell death. J Mol Cell Cardiol 2005; 38: 21–33.
Minamino T, Kitakaze M . ER stress in cardiovascular disease. J Mol Cell Cardiol 2010; 48: 1105–10.
Ma YM, Peng YM, Zhu QH, Gao AH, Chao B, He QJ, et al. Novel CHOP activator LGH00168 induces necroptosis in A549 human lung cancer cells via ROS-mediated ER stress and NF-kappaB inhibition. Acta Pharmacol Sin 2016; 37: 1381–90.
Travers KJ, Patil CK, Wodicka L, Lockhart DJ, Weissman JS, Walter P . Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation. Cell 2000; 101: 249–58.
Walter P, Ron D . The unfolded protein response: from stress pathway to homeostatic regulation. Science 2011; 334: 1081–6.
Beck D, Niessner H, Smalley KS, Flaherty K, Paraiso KH, Busch C, et al. Vemurafenib potently induces endoplasmic reticulum stress-mediated apoptosis in BRAFV600E melanoma cells. Sci Signal 2013; 6: ra7.
Liu MQ, Chen Z, Chen LX . Endoplasmic reticulum stress: a novel mechanism and therapeutic target for cardiovascular diseases. Acta Pharmacol Sin 2016; 37: 425–43.
Lv G, Lou Z, Chen S, Gu H, Shan L . Pharmacokinetics and tissue distribution of 2,3,5,4′-tetrahydroxystilbene-2-O-beta-D-glucoside from traditional Chinese medicine Polygonum multiflorum following oral administration to rats. J Ethnopharmacol 2011; 137: 449–56.
Zhou X, Yang Q, Xie Y, Sun J, Hu J, Qiu P, et al. Tetrahydroxystilbene glucoside extends mouse life span via upregulating neural klotho and downregulating neural insulin or insulin-like growth factor 1. Neurobiol Aging 2015; 36: 1462–70.
Zhang F, Wang YY, Yang J, Lu YF, Liu J, Shi JS . Tetrahydroxystilbene glucoside attenuates neuroinflammation through the inhibition of microglia activation. Oxid Med Cell Longev 2013; 2013: 680545.
Wang W, He Y, Lin P, Li Y, Sun R, Gu W, et al. In vitro effects of active components of Polygonum multiflorum Radix on enzymes involved in the lipid metabolism. J Ethnopharmacol 2014; 153: 763–70.
Xu XL, Huang YJ, Wang YQ, Chen XF, Zhang W . 2,3,4′,5-Tetrahydroxystilbene-2-O-beta-D-glucoside inhibits platelet-derived growth factor-induced proliferation of vascular smooth muscle cells by regulating the cell cycle. Clin Exp Pharmacol Physiol 2011; 38: 307–13.
Zhang W, Chen XF, Huang YJ, Chen QQ, Bao YJ, Zhu W . 2,3,4′,5-Tetrahydroxystilbene-2-O-beta-D-glucoside inhibits angiotensin II-induced cardiac fibroblast proliferation via suppression of the reactive oxygen species-extracellular signal-regulated kinase 1/2 pathway. Clin Exp Pharmacol Physiol 2012; 39: 429–37.
Xu XL, Zhu QY, Zhao C, Wang F, Zhou ZY, Hu YE, et al. The effect of 2,3,4′,5-tetrahydroxystilbene-2-O-beta-D-glucoside on pressure overload-induced cardiac remodeling in rats and its possible mechanism. Planta Med 2014; 80: 130–8.
Han X, Ling S, Gan W, Sun L, Duan J, Xu JW . 2,3,5,4′-tetrahydroxystilbene-2-O-beta-D-glucoside ameliorates vascular senescence and improves blood flow involving a mechanism of p53 deacetylation. Atherosclerosis 2012; 225: 76–82.
Hsu PL, Horng LY, Peng KY, Wu CL, Sung HC, Wu RT . Activation of mitochondrial function and Hb expression in non-haematopoietic cells by an EPO inducer ameliorates ischaemic diseases in mice. Br J Pharmacol 2013; 169: 1461–76.
Ye S, Tang L, Xu J, Liu Q, Wang J . Postconditioning's protection of THSG on cardiac ischemia-reperfusion injury and mechanism. J Huazhong Univ Sci Technolog Med Sci 2006; 26: 13–6.
Piazzi G, Bazzoli F, Ricciardiello L . Epigenetic silencing of Notch signaling in gastrointestinal cancers. Cell Cycle 2012; 11: 4323–7.
Marignol L, Rivera-Figueroa K, Lynch T, Hollywood D . Hypoxia, notch signalling, and prostate cancer. Nat Rev Urol 2013; 10: 405–13.
Nakagawa O, McFadden DG, Nakagawa M, Yanagisawa H, Hu T, Srivastava D, et al. Members of the HRT family of basic helix-loop-helix proteins act as transcriptional repressors downstream of Notch signaling. Proc Natl Acad Sci U S A 2000; 97: 13655–60.
MacGrogan D, Nus M, de la Pompa JL . Notch signaling in cardiac development and disease. Curr Top Dev Biol 2010; 92: 333–65.
Collesi C, Zentilin L, Sinagra G, Giacca M . Notch1 signaling stimulates proliferation of immature cardiomyocytes. J Cell Biol 2008; 183: 117–28.
Campa VM, Gutierrez-Lanza R, Cerignoli F, Diaz-Trelles R, Nelson B, Tsuji T, et al. Notch activates cell cycle reentry and progression in quiescent cardiomyocytes. J Cell Biol 2008; 183: 129–41.
Gude NA, Emmanuel G, Wu W, Cottage CT, Fischer K, Quijada P, et al. Activation of Notch-mediated protective signaling in the myocardium. Circ Res 2008; 102: 1025–35.
Wang X, Zhao L, Han T, Chen S, Wang J . Protective effects of 2,3,5,4′-tetrahydroxystilbene-2-O-beta-D-glucoside, an active component of Polygonum multiflorum Thunb, on experimental colitis in mice. Eur J Pharmacol 2008; 578: 339–48.
Chen T, Yang YJ, Li YK, Liu J, Wu PF, Wang F, et al. Chronic administration tetrahydroxystilbene glucoside promotes hippocampal memory and synaptic plasticity and activates ERKs, CaMKII and SIRT1/miR-134 in vivo. J Ethnopharmacol 2016; 190: 74–82.
Liu RR, Li J, Gong JY, Kuang F, Liu JY, Zhang YS, et al. MicroRNA-141 regulates the expression level of ICAM-1 on endothelium to decrease myocardial ischemia-reperfusion injury. Am J Physiol Heart Circ Physiol 2015; 309: H1303–13.
Boccalini G, Sassoli C, Formigli L, Bani D, Nistri S . Relaxin protects cardiac muscle cells from hypoxia/reoxygenation injury: involvement of the Notch-1 pathway. FASEB J 2015; 29: 239–49.
Yang Y, Duan W, Liang Z, Yi W, Yan J, Wang N, et al. Curcumin attenuates endothelial cell oxidative stress injury through Notch signaling inhibition. Cell Signal 2013; 25: 615–29.
Tao L, Wang Y, Gao E, Zhang H, Yuan Y, Lau WB, et al. Adiponectin: an indispensable molecule in rosiglitazone cardioprotection following myocardial infarction. Circ Res 2010; 106: 409–17.
Yang Y, Duan W, Lin Y, Yi W, Liang Z, Yan J, et al. SIRT1 activation by curcumin pretreatment attenuates mitochondrial oxidative damage induced by myocardial ischemia reperfusion injury. Free Radic Biol Med 2013; 65: 667–79.
Yu L, Sun Y, Cheng L, Jin Z, Yang Y, Zhai M, et al. Melatonin receptor-mediated protection against myocardial ischemia/reperfusion injury: role of SIRT1. J Pineal Res 2014; 57: 228–38.
Yang Y, Duan W, Jin Z, Yi W, Yan J, Zhang S, et al. JAK2/STAT3 activation by melatonin attenuates the mitochondrial oxidative damage induced by myocardial ischemia/reperfusion injury. J Pineal Res 2013; 55: 275–86.
Yang C, Wang Y, Liu H, Li N, Sun Y, Liu Z, et al. Ghrelin protects H9c2 cardiomyocytes from angiotensin II-induced apoptosis through the endoplasmic reticulum stress pathway. J Cardiovasc Pharmacol 2012; 59: 465–71.
Wu T, Dong Z, Geng J, Sun Y, Liu G, Kang W, et al. Valsartan protects against ER stress-induced myocardial apoptosis via CHOP/Puma signaling pathway in streptozotocin-induced diabetic rats. Eur J Pharm Sci 2011; 42: 496–502.
Zhang Z, Tong N, Gong Y, Qiu Q, Yin L, Lv X, et al. Valproate protects the retina from endoplasmic reticulum stress-induced apoptosis after ischemia-reperfusion injury. Neurosci Lett 2011; 504: 88–92.
Chen S, He FF, Wang H, Fang Z, Shao N, Tian XJ, et al. Calcium entry via TRPC6 mediates albumin overload-induced endoplasmic reticulum stress and apoptosis in podocytes. Cell Calcium 2011; 50: 523–9.
Griciuc A, Aron L, Ueffing M . ER stress in retinal degeneration: a target for rational therapy? Trends Mol Med 2011; 17: 442–51.
Bernales S, Papa FR, Walter P . Intracellular signaling by the unfolded protein response. Annu Rev Cell Dev Biol 2006; 22: 487–508.
Mera K, Kawahara K, Tada K, Kawai K, Hashiguchi T, Maruyama I, et al. ER signaling is activated to protect human HaCaT keratinocytes from ER stress induced by environmental doses of UVB. Biochem Biophys Res Commun 2010; 397: 350–4.
Harding HP, Zhang Y, Bertolotti A, Zeng H, Ron D . PERK is essential for translational regulation and cell survival during the unfolded protein response. Mol Cell 2000; 5: 897–904.
Marwarha G, Dasari B, Ghribi O . Endoplasmic reticulum stress-induced CHOP activation mediates the down-regulation of leptin in human neuroblastoma SH-SY5Y cells treated with the oxysterol 27-hydroxycholesterol. Cell Signal 2012; 24: 484–92.
Song W, Guo F, Zhong H, Liu L, Yang R, Wang Q, et al. Therapeutic window of globular adiponectin against cerebral ischemia in diabetic mice: the role of dynamic alteration of adiponectin/adiponectin receptor expression. Sci Rep 2015; 5: 17310.
Li H, Zhu X, Fang F, Jiang D, Tang L . Down-regulation of GRP78 enhances apoptosis via CHOP pathway in retinal ischemia-reperfusion injury. Neurosci Lett 2014; 575: 68–73.
Yu Y, Sun G, Luo Y, Wang M, Chen R, Zhang J, et al. Cardioprotective effects of notoginsenoside R1 against ischemia/reperfusion injuries by regulating oxidative stress- and endoplasmic reticulum stress- related signaling pathways. Sci Rep 2016; 6: 21730.
Kratsios P, Catela C, Salimova E, Huth M, Berno V, Rosenthal N, et al. Distinct roles for cell-autonomous Notch signaling in cardiomyocytes of the embryonic and adult heart. Circ Res 2010; 106: 559–72.
Ehebauer M, Hayward P, Martinez-Arias A . Notch signaling pathway. Sci STKE 2006; 2006: cm7.
Miele L . Notch signaling. Clin Cancer Res 2006; 12: 1074–9.
Croquelois A, Domenighetti AA, Nemir M, Lepore M, Rosenblatt-Velin N, Radtke F, et al. Control of the adaptive response of the heart to stress via the Notch1 receptor pathway. J Exp Med 2008; 205: 3173–85.
Urbanek K, Cabral-da-Silva MC, Ide-Iwata N, Maestroni S, Delucchi F, Zheng H, et al. Inhibition of notch1-dependent cardiomyogenesis leads to a dilated myopathy in the neonatal heart. Circ Res 2010; 107: 429–41.
Zhou XL, Wan L, Xu QR, Zhao Y, Liu JC . Notch signaling activation contributes to cardioprotection provided by ischemic preconditioning and postconditioning. J Transl Med 2013; 11: 251.
Pei H, Yu Q, Xue Q, Guo Y, Sun L, Hong Z, et al. Notch1 cardioprotection in myocardial ischemia/reperfusion involves reduction of oxidative/nitrative stress. Basic Res Cardiol 2013; 108: 373.
Rosati E, Sabatini R, De Falco F, Del Papa B, Falzetti F, Di Ianni M, et al. gamma-Secretase inhibitor I induces apoptosis in chronic lymphocytic leukemia cells by proteasome inhibition, endoplasmic reticulum stress increase and notch down-regulation. Int J Cancer 2013; 132: 1940–53.
Ohta K, Mizuno A, Li S, Itoh M, Ueda M, Ohta E, et al. Endoplasmic reticulum stress enhances gamma-secretase activity. Biochem Biophys Res Commun 2011; 416: 362–6.
Szabat M, Kalynyak TB, Lim GE, Chu KY, Yang YH, Asadi A, et al. Musashi expression in beta-cells coordinates insulin expression, apoptosis and proliferation in response to endoplasmic reticulum stress in diabetes. Cell Death Dis 2011; 2: e232.
Zhao J, Liang Y, Song F, Xu S, Nian L, Zhou X, et al. TSG attenuates LPC-induced endothelial cells inflammatory damage through notch signaling inhibition. IUBMB Life 2016; 68: 37–50.
Acknowledgements
This research was supported by the Major Science and Technology Projects “Innovative Drug Research Center of Shaanxi Province/Pharmaceutical Research of the TSG” in Shaanxi Province, China (2015SF2-08-01/S2016TNSF0060).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Zhang, M., Yu, Lm., Zhao, H. et al. 2,3,5,4′-Tetrahydroxystilbene-2-O-β-D-glucoside protects murine hearts against ischemia/reperfusion injury by activating Notch1/Hes1 signaling and attenuating endoplasmic reticulum stress. Acta Pharmacol Sin 38, 317–330 (2017). https://doi.org/10.1038/aps.2016.144
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/aps.2016.144
