
Abstract
Aim:
To investigate the roles of the calcineurin/nuclear factor of activated T cells (NFAT) pathway in regulation of wear particles-induced cytokine release and osteoclastogenesis from mouse bone marrow macrophages in vitro.
Methods:
Osteoclasts were induced from mouse bone marrow macrophages (BMMs) in the presence of 100 ng/mL receptor activator of NF-κB ligand (RANKL). Acridine orange staining and MTT assay were used to detect the cell viability. Osteoclastogenesis was determined using TRAP staining and RT-PCR. Bone pit resorption assay was used to examine osteoclast phenotype. The expression and cellular localization of NFATc1 were examined using RT-PCR and immunofluorescent staining. The production of TNFα was analyzed with ELISA.
Results:
Titanium (Ti) or polymethylmethacrylate (PMMA) particles (0.1 mg/mL) did not significantly change the viability of BMMs, but twice increased the differentiation of BMMs into mature osteoclasts, and markedly increased TNF-α production. The TNF-α level in the PMMA group was significantly higher than in the Ti group (96 h). The expression of NFATc1 was found in BMMs in the presence of the wear particles and RANKL. In bone pit resorption assay, the wear particles significantly increased the resorption area and total number of resorption pits in BMMs-seeded ivory slices. Addition of 11R-VIVIT peptide (a specific inhibitor of calcineurin-mediated NFAT activation, 2.0 μmol/L) did not significantly affect the viability of BMMs, but abolished almost all the wear particle-induced alterations in BMMs. Furthermore, VIVIT reduced TNF-α production much more efficiently in the PMMA group than in the Ti group (96 h).
Conclusion:
Calcineurin/NFAT pathway mediates wear particles-induced TNF-α release and osteoclastogenesis from BMMs. Blockade of this signaling pathway with VIVIT may provide a promising therapeutic modality for the treatment of periprosthetic osteolysis.
Similar content being viewed by others


Introduction
Total joint replacement (TJR) provides dramatic pain relief and joint function improvement for patients with end stage joint diseases. However, despite the popularity and initial success of TJR, periprosthetic osteolysis and subsequent aseptic loosening of prosthetic joint components remains a major problem for the long-term success and survival of prosthetic joints1. Although it is widely agreed that wear particles from the prosthesis play a central role in the initiation and development of osteolysis, the precise mechanism remains unclear2,3.
Substantial progress has been made in recent years in elucidating the pathophysiologic mechanisms responsible for periprosthetic osteolysis3. One area of investigation that has advanced rapidly is the demonstration that proinflammatory cytokines not only are produced in response to wear debris but also are responsible for the downstream processes leading to osteolysis3. Wear particles stimulate macrophages, foreign body giant cells, fibroblasts and T lymphocytes to produce a vast array of proinflammatory cytokines and other factors, including tumor necrosis factor-α (TNF-α), interleukin-1 (IL-1), IL-6, matrix metalloproteinases (MMPs) and peptides2,4,5. Of these factors, TNF-α is regarded as a key cytokine involved in the pathogenesis of aseptic loosening around implants6. TNF-α increases inflammation, induces differentiation of pre-osteoclasts, expands the osteoclast precursor pool, enhances osteoclast activation, and promotes osteoclast survival7,8. In addition, TNF-α augments expression of RANKL (receptor activator of NF-κB ligand) by osteoblasts and marrow stromal cells. RANKL is the key osteoclast differentiation factor, binding to RANK which is expressed on committed osteoclast precursors and promoting their differentiation into mature and functional osteoclasts9,10,11. Consequently, increased osteoclastogenesis results in unchecked bone erosion around the prosthetic implant9,12,13.
Osteoclasts are responsible for both physiological and pathological bone resorption14. Recently, it has been recognized that another transcription factor, nuclear factor of activated T cells (NFAT), functions downstream of RANKL signaling in osteoclast differentiation15,16. The NFAT transcription factor family comprises five members: NFATc1, NFATc2, NFATc3, NFATc4, and NFAT515. In resting cells, the NFAT members, except NFAT5, are retained in the cytoplasm by hyperphosphorylation of an N-terminal regulatory domain17. Signaling pathways culminating in a sustained calcium influx activate the phosphatase calcineurin, which dephosphorylates these NFAT proteins, promoting their nuclear translocation and activation17,18. Among NFAT members, NFATc1 plays a crucial role in osteoclastogenesis11. RANKL activates the TNF receptor-associated factor 6 (TRAF 6) and c-Fos pathways, leading to autoamplification of NFATc1, which mediates a gene expression program linked to cell fusion, osteoclast differentiation, and potentially to additional NFAT-mediated functions11,15,16,19.
Particulate wear debris can be generated in a variety of ways, and may include particles from all the various components of the orthopedic prosthesis (such as polyethylene, metal, and ceramic) as well as bone cement (polymethylmethacrylate, PMMA)20,21. In our previous study, we showed that inactivation of NFATc1 by 11R-VIVIT peptide, which is a specific peptide inhibitor of calcineurin-mediated NFAT activation18,22, potently blocked titanium (Ti) particle-induced osteoclastogenesis23. The cement-stem interface, however, represents a potential source of metal and PMMA particles. In the current work, we extended our study to investigate the effects of blocking the calcineurin/NFAT pathway on Ti and PMMA particle-induced proinflammatory cytokine release and osteoclastogenesis, and to demonstrate its therapeutic potential in treating or preventing inflammatory osteolysis.
Materials and methods
Reagents
Soluble recombinant mouse RANKL and macrophage colony-stimulating factor (M-CSF) were purchased from R&D Industries (Minneapolis, MN, USA). The 11R-VIVIT peptide (RRRRRRRRRRR-GGG-MAGPHPVIVITGPHEE) was purchased from Sigma Genosys (Woodlands, TX, USA). The mouse-specific enzyme-linked immunosorbent assay (ELISA) kits were purchased from Biosource International Inc (Camarillo, CA, USA).
Titanium and polymethylmethacrylate particle preparation
Commercial pure Ti particles, with an average diameter of 4.50 μm, were purchased from Johnson Matthey (Catalog #00681; Ward Hill, MA, USA) (Figure 1). Commercial PMMA particles, with an average diameter of 6.0 μm, were purchased from Polyscience Inc (Catalog #19130; Warrington, PA, USA) (Figure 1). The Ti particles were prepared as reported previously2,23. The PMMA particles were treated with 70% ethanol for 48 h to remove endotoxin24; after treatment the particles' endotoxin level was less than 0.1 EU/mL, as determined using a commercial detection kit (Chromogenic end-point TAL with a Diazo coupling kit, Xiamen Houshiji, Xiamen, China). Particle suspensions were sonicated for 30 min using an SB3200 Ultrasonic Generator (Shanghai Branson Ultrasonics Co Ltd, Shanghai, China) prior to incubation with cells. The concentration of particles used for incubation was 0.1 mg/mL. Such particles have been shown to effectively mimic wear particles retrieved from periprosthetic tissues9,25,26,27.

Scanning electron micrographs of titanium (Ti) and polymethylmethacrylate (PMMA) particles (×2000).
Animals and cell culture
Male C57BL/6J mice (5 weeks old), obtained from Shanghai SLAC laboratory Animal Co, Ltd (Shanghai, China) were used in this study. Approval was obtained from Shanghai Jiao Tong University School of Medicine Animal Studies Committee. Bone marrow macrophages (BMMs) were prepared from whole bone marrow as reported previously9,13,23,28. Briefly, bone marrow was flushed out from bilateral tibiae and femora and incubated in alpha minimal essential medium (α-MEM) containing 10% fetal bovine serum (FBS), 1% penicillin and streptomycin, and 10 ng/mL M-CSF. Cells were cultured in a 37 °C, 5% CO2 humidified incubator for 24 h. The non-adherent cells were collected and purified over a Histopaque gradient. Cells at the gradient interface were plated in α-MEM containing FBS and M-CSF and treated with or without 11R-VIVIT peptide (VIVIT) depending on the experimental design. M-CSF stimulated the gradient-purified cells to become BMMs. After 2 h, BMMs (1.5×106 cells/mL) were cultured in differentiation medium (DM) in the presence of soluble M-CSF and RANKL (30 ng/mL and 100 ng/mL, respectively) for 96 h, with or without the addition of Ti or PMMA particles (0.1 mg/mL). A 2.0 μmol/L concentration of VIVIT was added to cultures of BMMs pretreated with the same dose of VIVIT. Accordingly, there were six treatment groups of BMM cultures: 1) PMMA particles and VIVIT (PMMA/VIVIT); 2) Ti particles and VIVIT (Ti/VIVIT); 3) VIVIT only (VIVIT); 4) PMMA particles only (PMMA); 5) Ti particles only (Ti) and 6) neither wear particles nor VIVIT (control).
Acridine orange staining and MTT assay
Phagocytosis of wear particles was estimated by acridine orange staining as reported previously29. Briefly, BMMs (5×105 cells/mL) were cultured in M-CSF-containing α-MEM with or without Ti or PMMA particles for 24 and 48 h, and then fixed with 4% paraformaldehyde for 10 min. After permeabilizing with 0.1% Triton X-100 in buffer, cells were stained with 6 μg/mL acridine orange (Sigma Chemical Co, St Louis, MO, USA) in EDTA-buffer at room temperature for 10 min. Following washing with phosphate buffered saline (PBS), cells were covered with a 1:1 solution of glycerol and PBS to prevent photobleaching and observed using an inverted microscope (IX71S1F, Olympus, Tokyo, Japan) with a fluorescence attachment (BH2-RFL-T3, Olympus). The numbers of cells phagocytosing particles at different time points were quantified and expressed as a percentage of the total cells in the same visual field.
The effect of VIVIT and wear particles on the viability of BMMs was examined using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay23. BMMs (5×104 cells/well) were cultured in M-CSF-containing α-MEM with different treatments (see above) for 12, 24, and 48 h, and then incubated with 0.5 mg/mL MTT at 37 °C for 4 h. Following removal of supernatant, the insoluble formazan crystals were dissolved in 200 μL of dimethyl sulfoxide (DMSO) and absorbance was measured using a 570 nm wavelength Synergy HT microtiter plate reader (BioTek Instruments, Inc, VT, USA). Values were expressed as percent viability of the samples vs control cultures which were set as 100%29.
TRAP staining of osteoclasts
After 96 h, BMMs cultured in DM with different treatments were fixed and stained for tartrate-resistant acid phosphatase (TRAP) activity according to the manufacturer's instructions (Shanghai Rainbow Medical Reagent Research Co, Ltd, Shanghai, China). TRAP-positive cells with >3 nuclei were counted as osteoclasts12,23.
RNA isolation and reverse transcription polymerase chain reaction (RT-PCR)
After culture for 96 h, RT-PCR was used to assess the expression of NFATc1, TRAP, MMP-9, Cathepsin K (Cath-K), calcitonin receptor (CTR) and GAPDH mRNAs in BMMs following different treatments. Briefly, total RNA was isolated from BMMs using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. Each first-strand cDNA was synthesized by reverse transcription of 1 μg of total RNA with RevertAidTM M-MuLV Reverse Transcriptase (Fermentas, Glen Burnie, MD, USA). The cDNA was amplified using PCR; the primers and thermal cycling conditions for NFATc1, TRAP, Cath-K, CTR, and GAPDH were as reported previously23. For analysis of MMP-9 gene expression PCR amplification was performed using the following gene-specific primers: sense, 5′-CTGTCCAGACCAAGGGTACAGCCT-3′; antisense, 5′-GTGGTATAGTGGGACACATAGTGG-3′. Thermal cycling conditions were: 27 cycles of 94 °C for 45 s, 53 °C for 60 s, and 72 °C for 60 s. The amplified samples were run on a 1% agarose gel with ethidium bromide and the bands were visualized under UV illumination (Gel Documentation System; UVP, Upland, CA, USA). Values were normalized to GAPDH and analyzed by densitometry (BioRad, Hercules, CA, USA).
Bone pit resorption assay
BMMs were seeded directly onto ivory slices and cultured in DM with different treatments for 10 d, then removed with 1 mol/L NH4OH. The slices were mounted on a metal stub, gold coated, and examined using a JEOL scanning electron microscope (JSM-6360LV; JEOL Inc, Tokyo, Japan)23. The percentage area of resorption pits for each slice, the mean total number of pits formed, and the mean size of individual resorption lacunae (μm2) were quantified on SEM images captured from each quarter (×150 magnification) of the slice.
Measurement of proinflammatory cytokines by ELISA
BMMs under different treatment conditions were cultured in M-CSF-containing α-MEM for 6, 24, and 96 h. Cell culture supernatants were collected, centrifuged at 1200×g for 10 min and supplemented with a mixture of protease inhibitors (Kangchen, Shanghai, China). Mouse-specific ELISA kits were used to analyze the amounts of TNF-α produced by cells in accordance with the manufacturer's instructions. The intensity of color detected at 450 nm was measured. The sensitivity for TNF-α was 3 pg/mL.
Immunofluorescence staining of the NFATc1 protein
BMMs were cultured in DM with different treatments for 96 h. After incubation, cells were fixed in 4% paraformaldehyde for 20 min, treated with 0.2% Triton X-100 for 5 min and then sequentially incubated in 5% bovine serum albumin (BSA)/PBS for 30 min, mouse anti-NFATc1 monoclonal antibody (7A6; Santa Cruz, 2 μg/mL) for 60 min, and Alexa Fluor® 488-labeled anti mouse IgG antibody (Molecular Probes, Eugene, OR, USA, 4 μg/mL) for 60 min16,23. Cell nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI). Images were acquired using an inverted microscope (IX71S1F, Olympus, Tokyo, Japan) with a fluorescence attachment (BH2-RFL-T3, Olympus).
Statistical analysis
Data are expressed as mean±SEM. Differences between groups were analyzed using analysis of variance (ANOVA). Statistical significance was defined as P<0.05. Statistical analyses were performed using SPSS version 11.0 (SPSS, Chicago, IL, USA).
Results
Phagocytosis of wear particles by BMMs and the effects of VIVIT and wear particles on cell viability
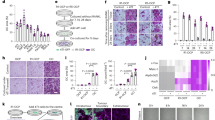
After culture with Ti or PMMA particles for 24 to 48 h, BMMs were found to phagocytose particles while retaining their normal morphology (Figure 2A). The number of cells phagocytosing particles increased markedly with prolonged culture time (Figure 2A and 2B). Viability of BMMs at each time-point was good, as shown in Figure 3. Compared to controls, treatment with 2.0 μmol/L VIVIT, addition of wear particles, or a combination of VIVIT and wear particles did not significantly reduce cell viability (Figure 3).

Phagocytosis of wear particles by BMMs. (A) BMMs challenged for 24 and 48 h with Ti and PMMA particles (acridine orange staining, ×20). (B) Numbers of cells phagocytosing particles (% of total cells in the same visual field) at different time intervals. Mean±SEM. n=12. cP<0.01 vs cells cultured with Ti particles for 24 h. fP<0.01 vs cells cultured with PMMA particles for 24 h.

The effects of VIVIT and wear particles on BMM viability. Mean±SEM. n=5. ODControl=0.1018 at 12 h; ODControl=0.1164 at 24 h; ODControl=0.1530 at 48 h.
Wear particles stimulate osteoclast formation and bone resorption
To determine the role of wear particles in osteoclast formation, we examined TRAP+ multinucleated cells (osteoclasts) in BMM culture in the presence of RANKL with or without Ti or PMMA particles. In the presence of RANKL, twice as many TRAP+ multinucleated cells were formed in cultures with Ti or PMMA particles than in those without particles (Figure 4A and 4B). In addition, more multinucleated cells were observed in the PMMA group than in the Ti group, although this difference was not significant. RT-PCR demonstrated that in the presence of RANKL the mRNAs of NFATc1, TRAP, MMP-9, Cath-K, and CTR were all expressed in BMM cultures both in the presence and the absence of wear particles (Figure 4C and 4D). However, the mRNA expression of each of these osteoclast-related genes was higher in cultures with particles than those without particles (Figure 4C and 4D). On the other hand, no osteoclasts were formed in cultures without RANKL (Figure 4A and 4B), and the osteoclast-related genes were only weakly expressed (Figure 4C and 4D).

Inhibition of wear particle-induced osteoclastogenesis by blockade of the calcineurin/NFAT pathway. (A) Osteoclasts differentiated from BMMs were identified by TRAP staining (×20). (B) Analysis of the number of osteoclasts formed from BMMs. (C) mRNA expression of NFATc1, TRAP, MMP-9, Cath-K, and CTR were determined by RT-PCR. (D) Relative mRNA expression of NFATc1, TRAP, MMP-9, Cath-K, and CTR in BMMs. Mean±SEM. n=5. bP<0.05, cP<0.01 vs cultures with RANKL alone. eP<0.05, fP<0.01 vs cultures with RANKL and Ti particles. hP<0.05, iP<0.01 vs cultures with RANKL and PMMA particles. ND: not detected.
To further determine the osteoclast phenotype of TRAP+ multinucleated cells induced by wear particles, we examined formation of resorption pits on ivory slices (Figure 5A). The results showed that the area resorbed on each slice and the total number of pits formed were both significantly greater on slices cultured with wear particles than those without particles (Figure 5B and 5C). However, there was no significant difference among these cultures in terms of the mean size of individual resorption lacunae (Figure 5D).

Inhibition of bone resorption by blockade of the calcineurin/NFAT pathway. (A) Complex resorption pits on ivory slices cultured with BMMs in the presence of RANKL (SEM, ×150). (B–D) Quantitative measurements of the percentage of resorption pit area of each slice (B), the mean total number of pits formed (C), and the mean size of individual resorption lacunae (D). Mean±SEM. n=12. cP<0.01 vs cultures with RANKL alone. fP<0.01 vs cultures with RANKL and Ti particles. iP<0.01 vs cultures with RANKL and PMMA particles.
Wear particles stimulate TNF-α production
The above experiments suggest that wear particles can strongly stimulate osteoclastogenesis, which is likely to be associated with increased levels of proinflammatory cytokines. Therefore, we monitored the concentration of TNF-α in BMM cultures using ELISA. After 6 h of culture, the concentration of TNF-α was significantly higher in the medium of cultures with wear particles than those without particles (Table 1). The concentrations of this cytokine increased further with culture time (Table 1). After 96 h of culture, the concentration of TNF-α in the PMMA group was significantly higher than that in cultures of cells with Ti particles (Table 1).
Expression of NFATc1 during wear particle-induced osteoclast differentiation
Although the NFAT transcription factor family was originally identified as controlling expression of cytokine and early immune-response genes in T cells, they are now found to regulate gene expression in a variety of nonimmune cells30. In this study, we monitored the expression of NFATc1 protein in BMMs and multinucleated cells cultured in RANKL-containing medium with or without Ti or PMMA particles. At 96 h, NFATc1 protein was strongly expressed in the nuclei of multinucleated cells (Figure 6), indicating nuclear translocation of NFATc1 during osteoclast differentiation. The numbers of TRAP+ multinucleated cells were much higher in cultures with wear particles than those without particles (Figure 6). On the other hand, when VIVIT was added, the expression of NFATc1 in BMMs was completely eliminated (Figure 6). Furthermore, RT-PCR also demonstrated that the expression of NFATc1 mRNA was also eliminated by VIVIT peptide (Figure 4C and 4D).

Expression of NFATc1 protein by BMMs and multinucleated cells in culture with different treatments (immunofluorescence staining and DAPI staining, ×40).
Effect of blocking the calcineurin/NFAT pathway on wear particle-induced osteoclast formation and bone resorption
Having demonstrated that NFATc1 is present in BMMs, we next wished to determine whether the suppression of NFATc1 activity affects wear particle-induced osteoclastogenesis. Our data demonstrated that VIVIT could profoundly inhibit osteoclastogenesis induced by RANKL in the presence or absence of wear particles (Figure 4A and 4B). RT-PCR showed that the expression of TRAP, MMP-9, Cath-K, and CTR genes were markedly inhibited by 2.0 μmol/L VIVIT after 96 h of culture with or without wear particles (Figure 4C and 4D). In addition, we found that VIVIT profoundly inhibited osteoclastic bone resorption, which was reflected by a significant reduction in total resorption area, total number of pits formed and the mean size of individual resorption lacunae (Figure 5A–5D).
Effect of blocking the calcineurin/NFAT pathway on wear particle-induced TNF-α release
To determine whether VIVIT can inhibit the production of proinflammatory cytokines induced by wear particles, we monitored the concentration of TNF-α in BMM culture using ELISA. When 2.0 μmol/L VIVIT was added to the culture medium together with wear particles, the concentration of TNF-α was significantly reduced (Table 1). Most importantly, after 96 h of culture, VIVIT reduced TNF-α production much more efficiently in the PMMA group than in the Ti group (P<0.01).
Discussion
To date the only established treatment for periprosthetic osteolysis and aseptic loosening is revision surgery, which is associated with greater morbidity and a poorer functional outcome31. Recent approaches to limiting aseptic loosening have focused on understanding and manipulating osteolysis at the molecular level through pharmacological interventions31.
Recent studies have revealed the biological cascade of events that are initiated by wear particles, mediate proinflammatory cytokine production and osteoclastogenesis, and finally result in bone loss around implants32. Osteolysis secondary to polyethylene debris is of paramount concern to today's joint replacement surgeons because significantly more polyethylene particles (70%–90%) appear in tissues around metal-on-polyethylene prostheses25,33. The cement-stem interface, however, represents a potential source of both metal and PMMA particles. These particles (metal and PMMA), though relatively less abundant in the tissue than polyethylene, are also an important factor contributing to osteolysis25,33,34,35. In this study, we utilized Ti and PMMA particles as a surrogate for wear debris to investigate the fundamental intracellular signaling mechanisms associated with implant particle-induced osteolysis.
The pathogenesis of osteolysis involves the activation of macrophages by wear debris and the release of various proinflammatory cytokines2,4,36. The exact mechanism of action and the specific role of each proinflammatory cytokine in this pathologic response are unclear, yet TNF-α has emerged as perhaps the critical cytokine28. Our results show that wear particles time-dependently induce TNF-α release by BMMs. However, consistent with previous studies37, TNF-α was unable to induce osteoclast maturation from precursors in the absence of RANKL. Nevertheless, while not necessary for osteoclast formation, TNF-α greatly enhances the differentiation of RANKL-primed osteoclast precursors, activation of mature osteoclasts, and osteoclast survival7. First and foremost, the production of TNF-α serves to stimulate recruitment of neutrophils and monocytes to the site of inflammation using two mechanisms: 1) stimulation of vascular cells to express new surface receptors and 2) stimulation of macrophages and other cells to secrete chemokines, including IL-1 and IL-638,39,40. In addition to direct actions on osteoclasts and their precursors, TNF-α may indirectly promote osteoclast activity through induction of expression of both RANKL and M-CSF in osteoblasts and marrow stromal cells21,36,41. Last but not least, TNF-α increases the expression of IP3R1, an important calcium channel responsible for evoking intracellular calcium oscillation, which promotes RANKL-induced osteoclastogenesis of mouse BMMs42. This scenario may explain why the increase in proinflammatory cytokines induced by prosthetic wear particles strongly enhances osteoclast formation and bone resorption in vivo.
Although it is generally accepted that interaction between particulate wear debris and macrophages is a critical initiating factor in periprosthetic osteolysis, there are still differences in the signaling properties among different types of wear particles27,34. In this study, we found that blockade of the calcineurin/NFAT pathway inhibited both Ti and PMMA particle-induced TNF-α release. Importantly, after 96 h of culture, this inhibitor reduced TNF-α production much more efficiently in the PMMA group than that in the Ti group. One possible mechanism is that NFAT participates in mediating the expression of proinflammatory cytokines induced by wear particles19,43. Blockade of NFAT by VIVIT would thus result in a reduction in the release of proinflammatory cytokines. Another possibility is that VIVIT might directly inhibit the expression of proinflammatory cytokines. However, further study is required to explore the different effects of VIVIT on the release of proinflammatory cytokines induced by different particles.
Osteoclastogenesis is the direct and central factor in the process of particulate wear debris-induced bone resorption and aseptic loosening6. The differentiation of osteoclast precursors into mature osteoclasts is regulated by two major cytokines: RANKL and M-CSF. M-CSF promotes the expansion and survival of osteoclast precursors, while RANKL initiates the signaling cascades leading to osteoclast differentiation17. In our study, TRAP+ multinucleated cells and resorption pits were both significantly increased in cultures of BMM with Ti and PMMA particles, in which the concentration of RANKL was the same as in controls. Without RANKL, however, wear particles were unable to induce differentiation of BMMs into osteoclasts. These data support the contention of Clohisy et al13 that RANKL is an essential cytokine mediating particle-induced osteoclast formation.
Recently, it has been demonstrated that RANKL promotes osteoclastogenesis through the induction and autoamplification of the key transcription factor NFATc111,15,16,19. Without activation of NFATc1, RANKL alone may not be sufficient to induce complete osteoclast differentiation44. Takayanagi et al16 reported that NFATc1−/− embryonic stem cells failed to differentiate into osteoclasts in response to RANKL stimulation. In contrast, the ectopic expression of NFATc1 could stimulate BMMs to undergo osteoclast differentiation even in the absence of RANKL stimulation16. These interesting data suggest that NFATc1 can induce osteoclastogenesis independent of RANKL. Compared to RANKL, however, much less is known about the effects of NFATc1 on implant particle-induced osteoclastogenesis.
Our study shows that NFATc1 is expressed by BMMs and multinucleated cells cultured with wear particles and RANKL. In the presence of RANKL, NFATc1 is expressed and translocated into the nucleus during osteoclast differentiation. More importantly, inactivation of NFATc1 by VIVIT results in a profound reduction in osteoclastogenesis despite the presence of RANKL and Ti or PMMA particles. Collectively, our data provide evidence that NFATc1 is essential for RANKL-induced osteoclast differentiation. Under these conditions, wear particles may increase the osteoclastogenic potential of BMMs. One can speculate that in vivo, these wear particles stimulate osteoclast formation, or osteoclastogenesis, leading to disruption of local bone structure.
NFATc1 is also expressed in mature osteoclasts, suggesting that the NFAT family is involved in control of osteoclast function. We found that osteoclastic bone resorption was significantly increased in the presence of wear particles. The cause of this stimulation is likely to be NFAT activation, which may not only accelerate osteoclastogenesis but also augment the bone-resorbing activity of each individual osteoclast. However, the mechanism by which NFATc1 affects osteoclast function remains unclear. It has been demonstrated that the final step of osteoclastogenesis is activation of resorption. To efficiently degrade mineralized tissue, osteoclasts secrete acid into a resorption lacuna formed between their apical membrane and the bone surface45. For each proton pumped into the lacuna, one bicarbonate ion remains in the cytoplasm45. To prevent cytoplasmic alkalinization, a basolateral bicarbonate/chloride exchanger, SLC4A2, provides egress for intracellular bicarbonate, which is induced during osteoclastogenesis through an NFATc1-dependent pathway45. Furthermore, in addition to the structural changes, the activity of osteoclasts is also associated with their viability. The increased resorption volume is consistent with the ability of RANKL and TNF-α to prolong the survival of osteoclasts7,46. Blockade of NFAT signal by VIVIT can suppress the increase in osteoclast survival induced by RANKL and TNF7,46.
The present study shows that blockade of the calcineurin/NFAT pathway by the VIVIT peptide potently impedes PMMA particle-induced osteoclastogenesis, which is in agreement with a previous report47. However, in this study, we provide evidence for the possible mechanisms by which blockade of the calcineurin/NFAT signaling cascade may be effective in preventing periprosthetic osteolysis. Our findings suggest that activation of the transcription factor NFATc1 appears to be central to both pathways induced by wear particles, ie proinflammatory cytokine release and osteoclastogenesis. Recently, Yarilina et al19 identified delayed TNF-induced signaling responses in primary human macrophages and found a sustained wave of signaling that begins after 1 to 2 d of TNF stimulation and is characterized by nuclear accumulation of NF-κB proteins and c-Jun and by calcium oscillations. These signaling events culminate in the induction and activation of NFATc119, which in turn regulates osteoclastogenesis and cytokine production, including TNF48,49. Blockade of the calcineurin/NFAT pathway with VIVIT may interrupt the positive feedback loop involving TNF and NFAT and offer a new opportunity for therapeutic intervention in TNF-driven chronic inflammatory diseases19.
All the above findings suggest that inactivation of NFATc1 may be a novel target for the treatment of wear particle-induced osteolysis. As the flow of information from calcium-mobilizing receptors to NFAT-dependent genes is critically dependent on interaction between the phosphatase calcineurin and the transcription factor NFAT18, a high-affinity calcineurin-binding peptide, VIVIT peptide, was developed based on the calcineurin docking motif of NFAT18. The peptide interferes with calcineurin-NFAT interaction, without compromising calcineurin phosphatase activity or non-NFAT-mediated signaling18,22. Thus, the VIVIT peptide constitutes a highly selective inhibitor of NFAT, which can be used to directly investigate the effect of NFAT on various cell types18. We found that VIVIT could significantly suppress the expression of NFATc1 in BMMs, which severely hampered osteoclastogenesis and cytokine release in response to wear particles. MTT assay showed that VIVIT did not compromise the viability of BMMs, suggesting that the decrease in osteoclastogenesis and TNF-α release is not due to a toxic effect.
In the present study, we investigated the role of the calcineurin/NFAT pathway in the regulation of proinflammatory cytokine release and osteoclastogenesis induced by wear particles. Our results indicate that Ti and PMMA particles, released during implant wear debris particles and aseptic loosening, constitute a major culprit by inducing inflammatory and osteolytic responses by macrophages and osteoclasts, respectively, and that activation of the transcription factor NFATc1 appears to be central to both proinflammatory cytokine release and osteoclastogenesis pathways induced by wear particles. Blockade of the calcineurin/NFAT pathway with VIVIT peptide may be a promising novel therapy not only for treating wear particle-induced periprosthetic osteolysis but also for controlling other chronic inflammatory bone diseases, such as rheumatoid arthritis and periodontal disease.
Author contribution
Feng-xiang LIU and Zhen-an ZHU designed the study; Feng-xiang LIU, Chuan-long WU, Mao-qiang LI, Yuan-qing MAO, Ming LIU, and Xiao-qing WANG performed the research; Feng-xiang LIU and De-gang YU participated in the data analysis and drafted the manuscript; Zhen-an ZHU and Ting-ting TANG revised the manuscript.
References
Yang SY, Wu B, Mayton L, Mukherjee P, Robbins PD, Evans CH, et al. Protective effects of IL-1Ra or vIL-10 gene transfer on a murine model of wear debris-induced osteolysis. Gene Ther 2004; 11: 483–91.
Rakshit DS, Ly K, Sengupta TK, Nestor BJ, Sculco TP, Ivashkiv LB, et al. Wear debris inhibition of anti-osteoclastogenic signaling by interleukin-6 and interferon-gamma. Mechanistic insights and implications for periprosthetic osteolysis. J Bone Joint Surg Am 2006; 88: 788–99.
Taki N, Tatro JM, Lowe R, Goldberg VM, Greenfield EM . Comparison of the roles of IL-1, IL-6, and TNFalpha in cell culture and murine models of aseptic loosening. Bone 2007; 40: 1276–83.
Holt G, Murnaghan C, Reilly J, Meek RM . The biology of aseptic osteolysis. Clin Orthop Relat Res 2007; 460: 240–52.
Holding CA, Findlay DM, Stamenkov R, Neale SD, Lucas H, Dharmapatni AS, et al. The correlation of RANK, RANKL and TNFalpha expression with bone loss volume and polyethylene wear debris around hip implants. Biomaterials 2006; 27: 5212–9.
Yang C, Niu S, Yu L, Zhu S, Zhu J, Zhu Q . The aqueous extract of Angelica sinensis, a popular Chinese herb, inhibits wear debris-induced inflammatory osteolysis in mice. J Surg Res 2012; 176: 476–83.
Nich C, Langlois J, Marchadier A, Vidal C, Cohen-Solal M, Petite H, et al. Oestrogen deficiency modulates particle-induced osteolysis. Arthritis Res Ther 2011; 13: R100.
Darowish M, Rahman R, Li P, Bukata SV, Gelinas J, Huang W, et al. Reduction of particle-induced osteolysis by interleukin-6 involves anti-inflammatory effect and inhibition of early osteoclast precursor differentiation. Bone 2009; 45: 661–8.
Clohisy JC, Hirayama T, Frazier E, Han SK, Abu-Amer Y . NF-kB signaling blockade abolishes implant particle-induced osteoclastogenesis. J Orthop Res 2004; 22: 13–20.
Nakashima T, Takayanagi H . The dynamic interplay between osteoclasts and the immune system. Arch Biochem Biophys 2008; 473: 166–71.
Asagiri M, Takayanagi H . The molecular understanding of osteoclast differentiation. Bone 2007; 40: 251–64.
Zheng H, Yu X, Collin-Osdoby P, Osdoby P . RANKL stimulates inducible nitric-oxide synthase expression and nitric oxide production in developing osteoclasts. An autocrine negative feedback mechanism triggered by RANKL-induced interferon-beta via NF-kappaB that restrains osteoclastogenesis and bone resorption. J Biol Chem 2006; 281: 15809–20.
Clohisy JC, Frazier E, Hirayama T, Abu-Amer Y . RANKL is an essential cytokine mediator of polymethylmethacrylate particle-induced osteoclastogenesis. J Orthop Res 2003; 21: 202–12.
Tsuji-Takechi K, Negishi-Koga T, Sumiya E, Kukita A, Kato S, Maeda T, et al. Stage-specific functions of leukemia/lymphoma-related factor (LRF) in the transcriptional control of osteoclast development. Proc Natl Acad Sci U S A 2012; 109: 2561–6.
Takayanagi H . The role of NFAT in osteoclast formation. Ann N Y Acad Sci 2007; 1116: 227–37.
Takayanagi H, Kim S, Koga T, Nishina H, Isshiki M, Yoshida H, et al. Induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of osteoclasts. Dev Cell 2002; 3: 889–901.
Aliprantis AO, Glimcher LH . NFATc1 in inflammatory and musculoskeletal conditions. Adv Exp Med Biol 2010; 658: 69–75.
Aramburu J, Yaffe MB, Lopez-Rodriguez C, Cantley LC, Hogan PG, Rao A . Affinity-driven peptide selection of an NFAT inhibitor more selective than cyclosporin A. Science 1999; 285: 2129–33.
Yarilina A, Xu K, Chen J, Ivashkiv LB . TNF activates calcium-nuclear factor of activated T cells (NFAT)c1 signaling pathways in human macrophages. Proc Natl Acad Sci U S A 2011; 108: 1573–8.
Saleh KJ, Thongtrangan I, Schwarz EM . Osteolysis: medical and surgical approaches. Clin Orthop Relat Res 2004: 138–47.
Purdue PE, Koulouvaris P, Nestor BJ, Sculco TP . The central role of wear debris in periprosthetic osteolysis. HSS J 2006; 2: 102–13.
Yu H, van Berkel TJ, Biessen EA . Therapeutic potential of VIVIT, a selective peptide inhibitor of nuclear factor of activated T cells, in cardiovascular disorders. Cardiovasc Drug Rev 2007; 25: 175–87.
Liu F, Zhu Z, Mao Y, Liu M, Tang T, Qiu S . Inhibition of titanium particle-induced osteoclastogenesis through inactivation of NFATc1 by VIVIT peptide. Biomaterials 2009; 30: 1756–62.
Koulouvaris P, Ly K, Ivashkiv LB, Bostrom MP, Nestor BJ, Sculco TP, et al. Expression profiling reveals alternative macrophage activation and impaired osteogenesis in periprosthetic osteolysis. J Orthop Res 2008; 26: 106–16.
von Knoch M, Jewison DE, Sibonga JD, Sprecher C, Morrey BF, Loer F, et al. The effectiveness of polyethylene versus titanium particles in inducing osteolysis in vivo. J Orthop Res 2004; 22: 237–43.
Lee SS, Woo CH, Chang JD, Kim JH . Roles of Rac and cytosolic phospholipase A2 in the intracellular signalling in response to titanium particles. Cell Signal 2003; 15: 339–45.
Rakshit DS, Lim JT, Ly K, Ivashkiv LB, Nestor BJ, Sculco TP, et al. Involvement of complement receptor 3 (CR3) and scavenger receptor in macrophage responses to wear debris. J Orthop Res 2006; 24: 2036–44.
Clohisy JC, Teitelbaum S, Chen S, Erdmann JM, Abu-Amer Y . Tumor necrosis factor-alpha mediates polymethylmethacrylate particle-induced NF-kappaB activation in osteoclast precursor cells. J Orthop Res 2002; 20: 174–81.
Ciapetti G, Gonzalez-Carrasco JL, Savarino L, Montealegre MA, Pagani S, Baldini N . Quantitative assessment of the response of osteoblast- and macrophage-like cells to particles of Ni-free Fe-base alloys. Biomaterials 2005; 26: 849–59.
Huang H, Chikazu D, Voznesensky OS, Herschman HR, Kream BE, Drissi H, et al. Parathyroid hormone induction of cyclooxygenase-2 in murine osteoblasts: role of the calcium-calcineurin-NFAT pathway. J Bone Miner Res 2010; 25: 819–29.
Shin DK, Kim MH, Lee SH, Kim TH, Kim SY . Inhibitory effects of luteolin on titanium particle-induced osteolysis in a mouse model. Acta Biomater 2012; 8: 3524–31.
Liu YP, Yu GR, Li K, Yuan F . Is there another possible approach to inhibit wear particles-induced inflammatory osteolysis? Med hypotheses 2011; 76: 280–2.
Baumann B, Rader CP, Seufert J, Noth U, Rolf O, Eulert J, et al. Effects of polyethylene and TiAlV wear particles on expression of RANK, RANKL and OPG mRNA. Acta Orthop Scand 2004; 75: 295–302.
Masui T, Sakano S, Hasegawa Y, Warashina H, Ishiguro N . Expression of inflammatory cytokines, RANKL and OPG induced by titanium, cobalt-chromium and polyethylene particles. Biomaterials 2005; 26: 1695–702.
Yamanaka Y, Abu-Amer Y, Faccio R, Clohisy JC . Map kinase c-JUN N-terminal kinase mediates PMMA induction of osteoclasts. J Orthop Res 2006; 24: 1349–57.
Jin S, Park JY, Hong JM, Kim TH, Shin HI, Park EK, et al. Inhibitory effect of (–)-epigallocatechin gallate on titanium particle-induced TNF-alpha release and in vivo osteolysis. Exp Mol Med 2011; 43: 411–8.
Lam J, Takeshita S, Barker JE, Kanagawa O, Ross FP, Teitelbaum SL . TNF-alpha induces osteoclastogenesis by direct stimulation of macrophages exposed to permissive levels of RANK ligand. J Clin Invest 2000; 106: 1481–8.
Wei S, Kitaura H, Zhou P, Ross FP, Teitelbaum SL . IL-1 mediates TNF-induced osteoclastogenesis. J Clin Invest 2005; 115: 282–90.
Suarez-Cuervo C, Harris KW, Kallman L, Vaananen HK, Selander KS . Tumor necrosis factor-alpha induces interleukin-6 production via extracellular-regulated kinase 1 activation in breast cancer cells. Breast Cancer Res Treat 2003; 80: 71–8.
Cunningham BW, Orbegoso CM, Dmitriev AE, Hallab NJ, Sefter JC, Asdourian P, et al. The effect of spinal instrumentation particulate wear debris. An in vivo rabbit model and applied clinical study of retrieved instrumentation cases. Spine J 2003; 3: 19–32.
Datta HK, Ng WF, Walker JA, Tuck SP, Varanasi SS . The cell biology of bone metabolism. J Clin Pathol 2008; 61: 577–87.
Xia L, Zhang D, Wang C, Wei F, Hu Y . PC-PLC is involved in osteoclastogenesis induced by TNF-alpha through upregulating IP3R1 expression. FEBS Lett 2012; 586: 3341–8.
Minematsu H, Shin MJ, Celil Aydemir AB, Seo SW, Kim DW, Blaine TA, et al. Orthopedic implant particle-induced tumor necrosis factor-alpha production in macrophage-monocyte lineage cells is mediated by nuclear factor of activated T cells. Ann N Y Acad Sci 2007; 1117: 143–50.
Ishida N, Hayashi K, Hoshijima M, Ogawa T, Koga S, Miyatake Y, et al. Large scale gene expression analysis of osteoclastogenesis in vitro and elucidation of NFAT2 as a key regulator. J Biol Chem 2002; 277: 41147–56.
Wu J, Glimcher LH, Aliprantis AO . HCO3−/Cl− anion exchanger SLC4A2 is required for proper osteoclast differentiation and function. Proc Natl Acad Sci U S A 2008; 105: 16934–9.
Pereverzev A, Komarova SV, Korcok J, Armstrong S, Tremblay GB, Dixon SJ, et al. Extracellular acidification enhances osteoclast survival through an NFAT-independent, protein kinase C-dependent pathway. Bone 2008; 42: 150–61.
Yamanaka Y, Abu-Amer W, Foglia D, Otero J, Clohisy JC, Abu-Amer Y . NFAT2 is an essential mediator of orthopedic particle-induced osteoclastogenesis. J Orthop Res 2008; 26: 1577–84.
Canellada A, Cano E, Sanchez-Ruiloba L, Zafra F, Redondo JM . Calcium-dependent expression of TNF-alpha in neural cells is mediated by the calcineurin/NFAT pathway. Mol Cell Neurosci 2006; 31: 692–701.
Kaminuma O, Kitamura F, Kitamura N, Hiroi T, Miyoshi H, Miyawaki A, et al. Differential contribution of NFATc2 and NFATc1 to TNF-alpha gene expression in T cells. J Immunol 2008; 180: 319–26.
Acknowledgements
This work was supported by grants from the National Natural Science Foundation of China (No 81171729), the Fund for Key Disciplines of Shanghai Municipal Education Commission (J50206) and the Program for the Shanghai Key Laboratory of Orthopedic Implants (No 08DZ2230330).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Liu, Fx., Wu, Cl., Zhu, Za. et al. Calcineurin/NFAT pathway mediates wear particle-induced TNF-α release and osteoclastogenesis from mice bone marrow macrophages in vitro. Acta Pharmacol Sin 34, 1457–1466 (2013). https://doi.org/10.1038/aps.2013.99
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/aps.2013.99
Keywords
This article is cited by
-
Progranulin suppresses titanium particle induced inflammatory osteolysis by targeting TNFα signaling
Scientific Reports (2016)
-
Traffic-Related PM2.5 Induces Cytosolic [Ca2+] Increase Regulated by Orai1, Alters the CaN-NFAT Signaling Pathway, and Affects IL-2 and TNF-α Cytoplasmic Levels in Jurkat T-Cells
Archives of Environmental Contamination and Toxicology (2015)
