
Abstract
Aim:
To investigate the effects of curcumin on proliferation and apoptosis in testicular cancer cells in vitro and to investigate its molecular mechanisms of action.
Methods:
NTera-2 human malignant testicular germ cell line and F9 mouse teratocarcinoma stem cell line were used. The anti-proliferative effect was examined using MTT and colony formation assays. Hoechst 33258 staining, TUNEL and Annexin V-FITC/PI staining assays were used to analyze cell apoptosis. Protein expression was examined with Western blot analysis and immunocytochemical staining.
Results:
Curcumin (5, 10 and 15 μmol/L) inhibited the viability of NTera-2 cells in dose- and time-dependent manners. Curcumin significantly inhibited the colony formation in both NTera-2 and F9 cells. Curcumin dose-dependently induced apoptosis of NTera-2 cells by reducing FasL expression and Bcl-2-to-Bax ratio, and activating caspase-9, -8 and -3. Furthermore, curcumin dose-dependently reduced the expression of AP transcription factor AP-2γ in NTera-2 cells, whereas the pretreatment with the proteasome inhibitor MG132 blocked both the curcumin-induced reduction of AP-2γ and antiproliferative effect. Curcumin inhibited ErbB2 expression, and decreased the phosphorylation of Akt and ERK in NTera-2 cells.
Conclusion:
Curcumin induces apoptosis and inhibits proliferation in NTera-2 cells via the inhibition of AP-2γ-mediated downstream cell survival signaling pathways.
Similar content being viewed by others


Introduction
Testicular germ cell tumors (TGCTs), a complex tumor type that is composed of several different pathogenesis, pathological types and clinical manifestations, account for 90% of testicular cancer cases. The overall incidence of TGCT increased from 3.35 per 100 000 to 4.84 per 100 000 men from 1973 to 19981. Treatment strategies for TGCTs include a combination of surgery, radiotherapy and chemotherapy. Unfortunately, while some patients are eligible for curative treatment, recurrence is a frequent issue for many patients after tumor ablation2. Accordingly, an urgent need exists to identify new therapeutic agents for the treatment of TGCTs in clinical practice.
Natural products occupy a very important position in the area of cancer chemotherapy due to their excellent pharmacological activities and low toxicity. Curcumin, a type of yellow pigment that is extracted from the rhizome of turmeric, has been used for centuries as a food additive and a traditional medicine in Asian countries3. Recently, curcumin gained attention as an anti-cancer agent due to its chemopreventive and chemotherapeutic potential, while having no discernable side effects4,5. The molecular mechanisms underlying the anticancer effects of curcumin have been studied in some detail, but are not completely understood. Curcumin inhibits proliferation in a variety of cancer cells through targeting multiple cellular signaling pathways, including the nuclear factor kappa B, Wnt, mitogen-activated protein kinase (MAPK), and Notch signaling pathways6,7. Curcumin has a diverse range of molecular targets, including cyclooxygenase-2 (COX-2), protein kinase C, thioredoxin reductase, 5-lipoxygenase (5-LOX), and tubulin, which supports the concept that it may act upon numerous biochemical and molecular cascades8. Studies have shown that curcumin inhibits cell proliferation and induces apoptosis in many cancers, such as breast cancer, gastric cancer, prostate cancer, colon cancer and melanoma9,10.
AP-2γ is a member of the AP transcription factor family, which plays an important role in the development and differentiation of trophoblasts and germ cells. AP-2γ is expressed in early gonocytes, but it is not present in any other germ cells in the prepubertal or postpubertal testes. Recent reports demonstrated that AP-2γ is not only involved in self-renewal, but is also specifically responsible for the survival of immature germ cells and tissue-specific stem cells; these findings suggest that AP-2γ functions as a master switch during the differentiation of human testicular germ cell tumors11,12. Interestingly, AP-2γ is also expressed in ovarian cancer, breast cancer, squamous cell carcinoma, and prostate cancer13,14,15, but little is known about its potential function in malignant disease. Over-expression of AP-2γ has been correlated with enhanced tumorigenicity, enhanced metastatic potential and poor prognosis. Moreover, AP-2γ over-expression increases the malignant potential of tumors, whereas AP-2γ down-regulation inhibits tumor growth, suggesting that AP-2γ might play a key role in maintaining cancer cell survival16,17. Altogether, these findings suggest that AP-2 participates in complex biological processes, including cell apoptosis and tumor formation. Recent studies demonstrated that AP-2γ is a prognostic factor in human TGCTs, and it is currently considered to be a target for TGCT therapy18.
The aims of this study were to determine effects of curcumin on proliferation and apoptosis in NTera-2 cells and to investigate its molecular mechanisms of action.
Materials and methods
Reagents and cell culture
Curcumin, from curcuma longa crystalline, was purchased from the Sigma-Aldrich Trading Co, Ltd (Shanghai, China), and it was dissolved in DMSO to obtain a final concentration of 1 mmol/L. Then, it was kept at -20 °C until use. The anti-caspase 3, 8, 9, anti-PARP, anti-AP-2γ, anti-ErbB2, and anti-GAPDH antibodies were purchased from Santa Cruz (Santa Cruz, CA, USA). Anti-ERK, pERK, AKT, and pAKT antibodies were purchased from Cell Signaling Technology Inc (Trask Lane, Danvers, MA, USA). Anti-rabbit and anti-mouse secondary antibodies for Western blotting or immunofluoresence staining were purchased from Santa Cruz (Santa Cruz, USA). The F9 mouse teratocarcinoma stem cell line and the NTera-2 human malignant testicular germ cell line were obtained from American Tissue Culture Collection (ATCC, Manassas, VA, USA). F9 cells were cultured in DMEM medium (Gibco) supplemented with 10% HyClone fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin at 37 °C with 5% CO2. NTera-2 cells were cultured in MEM Alpha medium (Gibco) supplemented with 10% Fetal Bovine Serum (Gibco BRL), 100 U/mL penicillin, and 100 μg/mL streptomycin at 37 °C, 5% CO2.
Cell viability assay
The effect of curcumin on NTera-2 cell proliferation was assessed using MTT and colony formation assays. NTera-2 cells were maintained in MEM Alpha Medium until the mid-log phase, and they were then seeded in 24-well plates at a density of 80%–90% confluence, with or without various concentrations of curcumin (5, 10, and 15 μmol/L). One hundred microliters of MTT was added to each well, and the cells were incubated at 37 °C for 4 h. After removing the supernatant, 300 μL of DMSO was added to dissolve the formazan crystals, and the optical density was detected at 490 nm. The data represent the mean of three readings, and each dose was tested in triplicate.
We detected the effect of curcumin on proliferation in a variety of cultured cell lines (NTera-2 and F-9). Briefly, 6-well plates were seeded with 500 viable cells, and they were allowed to grow for 24 h. The cells were then incubated in the presence or absence of 10 μmol/L curcumin for 24 h. The curcumin containing medium was then removed, and the cells were washed in PBS and incubated for an additional 15 d in complete medium. Each treatment was performed in triplicate. The resulting colonies were washed twice with PBS and fixed in methanol for 15 min at room temperature, followed by staining with 20% Giemsa solution for 30 min. The colonies were counted and compared with untreated cells.
Morphological evaluation of apoptotic cells
NTera-2 cells were treated with 10 μmol/L curcumin for 24 h, and nuclear fragmentation was visualized by Hoechst 33258 staining. Briefly, 1×105 cells were seeded in 12-well plates, and they were incubated with curcumin. After 24 h, the cells were collected, washed twice with PBS and fixed in 4% paraformaldehyde for 10 min at room temperature before being deposited on polylysine-coated slides. After 30 min, the adherent cells were incubated with 0. 1% Triton X-100 for 5 min at 4 °C. The cells were then incubated with 50 μmol/L Hoechst 33258 solution for 10 min at room temperature. After washing three times with PBS, the cells were viewed under a fluorescence microscope.
Flow cytometric analysis
Cells were incubated for 24 h with different concentration of curcumin (0, 5, 10, and 15 μmol/L). Apoptosis was determined by flow cytometry using the Annexin V-FITC/propidium iodide (PI) protocol described by the manufacturer (Beckman Coulter)19.
TUNEL assay
Apoptotic endonucleases can generate free 3'-OH groups at the ends DNA fragments, which can be identified by TUNEL staining and used for the detection of apoptosis. NTera-2 cells were cultured in 12-well plates (1×105 cells/well) covered with the appropriate slips for 12 h. Then, curcumin (5, 10, 15, 20, and 25 μmol/L) was added to the fresh medium. Twenty-four hours later, the cells were washed with cold PBS and fixed with 4% paraformaldehyde. TUNEL assays were performed using the Colorimetric TUNEL Apoptosis Assay Kit, according to the manufacturer's instructions. Briefly, biotinylated nucleotides were added to DNA breaks with free 3'-hydroxyl residues by terminal deoxynucleotidyl transferase. Labeled DNA was then identified using avidin peroxidase, using diaminobenzidine as a substrate. Apoptotic cells displayed brown staining in the nucleus.
Western blot analysis
Treated cells were harvested at the indicated points and lysed in RIPA buffer containing a protease inhibitor and phosphatase inhibitor cocktail. After 3 freeze-thaw cycles in liquid nitrogen, the resulting cell lysates were cleared by centrifugation at 12 000×g for 10 min at 4 °C, and the proteins were separated by 10% SDS-PAGE. After electrophoresis, the proteins were transferred to polyvinylidene difluoride-plus membranes. The membranes were blocked with 50 g/L nonfat milk in PBST washing buffer (PBS, 0.05% Tween-20) for 30 min, and then incubated with the indicated primary antibodies at 1:1000 (Bcl-2, Bax, Cyt c, FasL, caspase-3, caspase-8, caspase-9, PARP, AP-2γ, ErbB2, ERK, pERK, AKT, pAKT, GAPDH, and β-actin) for 3 h or overnight at 4 °C. Then, the membranes were incubated with a 1:2000 dilution of the proper ALP-conjugated secondary antibody for 1 h at room temperature. After four washes, the protein signals were visualized using an ECL Plus Kit, according to the manufacturer's instructions. The experiments were repeated at least twice, with protein extracts harvested independently. Densitometry was performed using ImageJ software.
RNA extraction and RT-PCR
NTera-2 cells were incubated with different doses of curcumin for 24 h. Total RNA was extracted using the TRIzol reagent (Invitrogen, CA, USA), according to the manufacturer's instructions. For RT-PCR, 2 μg of total RNA was subjected to a reverse transcription step using Promega reagents (USA). For each sample, mRNA levels of the target genes were corrected for GAPDH mRNA levels. The following PCR primers were used: AP-2γ sense: 5′-ATCTTGGAGGACGAAATGAGAT-3′, anti-sense: 5′-CAGATGGCCTGGCTGCCAA-3′; GAPDH: sense: 5′-GACTGTCTCCTCCCAAATTT-3′, anti-sense: 5′-GCATGGACTGTGGTCATGAGT-3′.
Transient transfection
NTera-2 cells were grown to 85% confluence in 100 mm culture dishes and transfected with 600 pmol of AP-2γ siRNA or scramble siRNA. Twenty-four hours after transfection, the cells were collected, and total protein lysates were prepared for immunoblot analysis. The siRNAs (Shanghai GenePharma) were as follows: AP-2γ siRNA (sense), 5′-GCUCUACGUCUAAAUACAATT-3′; AP-2γ siRNA (antisense), 5′-UUGUAUUUAGACGUAGAGCTG-3′; scramble siRNA (sense), 5′-UUCUCCGAACGUGUCACGUTT-3′; scramble siRNA (antisense), 5′-ACGUGACACGUUCGGAGAATT-3′.
Immunocytochemical staining
To examine the role of the proteasome in the effect of curcumin, we pre-incubated NTera-2 cells with the proteasome inhibitor, MG132 (100 μmol/L), for 5 h prior to the addition of curcumin (10 μmol/L) for 24 h. Cells were washed twice in PBS for 10 min, then fixed with 4% (w/v) paraformaldehyde in PBS for 10 min. Cells were incubated with anti-AP-2γ monoclonal primary antibody diluted in PBS with 10% (v/v) normal goat serum for 1 h. Then, the cells were incubated with the Alexa Red-conjugated anti-mouse IgG (Molecular Probes) secondary antibody under the same conditions. Hoechst 33258 was used to stain the nuclei.
Statistical analysis
Each experiment was repeated at least three times. All data are presented as the mean±SD. Differences between the control and treated groups were assessed using an unpaired Student's t-test. A P<0.05 was considered to be statistically significant.
Results
Curcumin treatment inhibited the proliferation of testicular germ cell lines
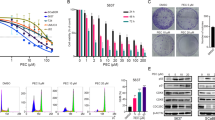
The MTT assay was used to assess the cytotoxicity of different concentrations of curcumin (0, 5, 10, and 15 μmol/L) on NTera-2 cells after 8, 16, 24, and 32 h. As shown in Figure 1A, there was approximately a 3-fold decrease in cell survival between the control (0 μmol/L) and curcumin (15 μmol/L) treated cells at 32 h. In addition, at a 15 μmol/L dose, curcumin decreased cell proliferation by approximately 2.5-fold between 0 and 32 h. The IC50 values for curcumin in NTera-2 cells at 8, 16, 24, and 32 h were 12.46, 16.97, 10.70, and 8.87 μmol/L, respectively. These results indicate that curcumin inhibits the growth of NTera-2 cells in a dose- and time-dependent manner (P<0.05). To determine the long-term effect of curcumin treatment, cells were treated with 10 μmol/L curcumin for 24 h, and then the cells were allowed to grow in normal media. Curcumin treatment suppressed colony formation in all testicular germ cell lines tested (Figure 1B), suggesting that curcumin's effect on the tumor cells was irreversible. The numbers of colonies formed in the presence of curcumin (10 μmol/L) dropped to 64.14% and 66.25% in NTera-2 and in F9 cells, respectively, compared to the controls.

Curcumin treatment inhibited the proliferation of testicular germ cell lines. (A) Curcumin inhibited the viability of NTera-2 cells at 8, 16, 24, and 32 h, in a time- and dose-dependent manner. bP<0.05, cP<0.01 vs control group. (B) Cell viability was determined using colony formation assays. Testicular cancer cells were incubated with curcumin (10 μmol/L) for 24 h, and then the cells were allowed to grow into colonies for 15 d. Incubation with curcumin inhibits colony formation. Data represent the mean±SD of three independent experiments. cP<0.01 vs control cells using the Student's t-test.
Curcumin treatment induced apoptosis in NTera-2 cells
Hoechst staining revealed typical morphological changes, such as the formation of apoptotic bodies, after 24 h treatment with 10 μmol/L curcumin, whereas the control cells did not show apoptosis-related morphological changes (Figure 2A). Normal nuclei were identified as having non-condensed chromatin dispersed over the entire nucleus, and apoptotic nuclei were identified as having condensed chromatin that was contiguous with the nuclear membrane and/or fragmented nuclei. As shown in Figure 2B, TUNEL staining was detected in NTera-2 cells after curcumin treatment. TUNEL-positive (brown staining) cells were characterized as apoptotic cells. The brown signal significantly increased with increasing concentrations of curcumin. Annexin V-FITC/PI double-labeled flow cytometry was used to assess the ratio of apoptotic NTera-2 cells following curcumin treatment. The total apoptosis ratio was the sum of the early apoptotic and late apoptotic ratios. The apoptosis rates for NTera-2 cells treated with 5, 10, and 15 μmol/L curcumin for 24 h were 7.8%±0.58%, 13.0%±1.06%, and 18%±1.78%, respectively, which were significantly higher than that of the control group (4.6%±0.31%) (Figure 2C). Altogether, these results provide substantial evidence that curcumin induces apoptosis in NTera-2 cells.

Effects of curcumin on apoptosis in NTera-2 cells. (A) The morphology of apoptotic nuclei was observed after Hoechst staining using a fluorescence microscope (magnification ×40). The control group was treated with DMSO. (B) A TUNEL Apoptosis Assay Kit was used to detect apoptosis in NTera-2 cells. The brown staining, indicative of apoptotic nuclei, significantly increased with increasing concentration of curcumin. (C) The apoptosis rate was analyzed by flow cytometry. Numbers in the respective quadrant profiles indicate the percentage of apoptotic cells. Each bar corresponds to the mean±SD of three independent experiments. bP<0.05, cP<0.01 vs control cells.
Curcumin-induced apoptosis is mediated via caspase activation in NTera-2 cells
Caspases are cell-death proteases that play a significant role in both the initiation and execution of apoptotic programs in diverse species20,21. Caspase assays revealed that curcumin induced caspase 9, 8, and 3 activation in a dose-dependent manner, and the levels of cleaved PARP also increased (Figure 3B). Western blot analyses of the anti-apoptotic proteins, Bcl-2 and FasL and the pro-apoptotic protein, Bax, confirmed that curcumin treatment caused apoptosis. Curcumin treatment inhibited the expression of Bcl-2 and FasL and increased Bax protein levels (Figure 3A). These data suggest that curcumin is a potent inducer of apoptosis in malignant testicular germ cells.

Curcumin modulates the expression of apoptotic genes in NTera- 2 cells. Cells were treated with different concentrations of curcumin for 24 h, as indicated. (A) Curcumin reduced the expression of the anti-apoptotic proteins, Bcl-2 and FasL, whereas it increased the expression of pro-apoptotic proteins compared to untreated cells. (B) Western blotting analyses showed a reduction of cytochrome c and the cleavage of caspase-9, -8, -3, and PARP in response to various concentrations of curcumin. β-actin was used as a control. One representative of three different experiments is shown.
Curcumin blocks the amplification of AP-2γ and reduces its expression in a dose-dependent manner
AP-2γ was reported to be a marker of testicular germ cell tumors. Consequently, drugs that can deregulate the expression of AP-2γ could potentially have therapeutic value. RT-PCR analysis and Western blot analyses showed that curcumin suppressed AP-2γ amplification and reduced AP-2γ expression in NTera-2 cells. The most significant decrease in AP-2γ expression was observed in response to 15 μmol/L of curcumin (Figure 4A).

The effect of siRNA-AP-2γ was consistent with that of curcumin treated in NTera-2 cells. (A) NTera-2 cells were treated with indicated dose of curcumin. RT-PCR and Western blot assays were used to monitor AP-2γ expression. Expression was normalized to GAPDH. (B) Western blot analysis of AP-2γ expression, cleaved PARP, caspase-8, caspase-9, caspase-3, and β-actin after 24 h treatment with siRNA-AP-2γ or siRNA-NC.
The effect of AP-2γ siRNA correlated with that of curcumin treatment in NTera-2 cells
To determine whether AP-2γ can regulate apoptosis in NTera-2 cells, we conducted Western blotting assays. These analyses revealed the cleavage of apical pro-caspase-3, pro-caspase-8, pro-caspase-9, and PARP into their active fragments in NTera-2 cells transfected with AP-2γ siRNA (Figure 4B). These data suggest that curcumin produces the same effect as AP-2γ knockdown in NTera-2 cells and that curcumin can regulate apoptosis of NTera-2 cell by targeting AP-2γ.
Curcumin induces the proteasome-mediated degradation of AP-2γ
To elucidate the mechanism by which curcumin inhibits the transcriptional activity of AP-2γ, we examined whether curcumin affected the level of AP-2γ protein degradation by the ubiquitin-proteasome pathway. As shown in Figure 5A, in the presence of a proteasome inhibitor, MG132, the ability of curcumin to inhibit AP-2γ was blocked. This result was confirmed by immunocytochemical staining of AP-2γ expression in NTera-2 cells (Figure 5B). MG132 also blocked the anti-proliferative effect of curcumin in the same conditions (Figure 5C). Thus, our results indicated that curcumin treatment promotes the proteasomal degradation of AP-2γ.

The proteasome inhibitor, MG132, blocks the curcumin-mediated reduction of AP-2γ. (A) Anti-AP-2γ Western blot of whole cell extracts incubated with the MG132 (100 μmol/L) for 5 h, prior to curcumin (10 μmol/L) treatment for 24 h. (B) Immunocytochemical staining of AP-2γ expression. (C) MG132 reversed the curcumin-mediated inhibition of cell proliferation, as determined by MTT assays. Data represent the mean±SD of three independent experiments. cP<0.01 vs curcumin.
Detection of ErbB2, ERK, and AKT
To assess whether blocking AP-2γ with curcumin affects its downstream signaling pathways, whole cell protein extracts were analyzed by Western blotting. When treated with curcumin, the phosphorylation of Akt and ERK decreased, and ErbB2 was also down-regulated in a dose-dependent manner in NTera-2 cells. No decrease in total ERK and AKT protein levels was observed (Figure 6). Therefore, curcumin can significantly reduce the activation of the ERK and AKT kinases.

Effect of curcumin on the expression of ErbB2 and the phosphorylation of ERK and AKT in NTera-2 cells. β-actin was used as an internal standard. Bands were analyzed using ImageJ software. The values represent the mean±SD of three assays. bP<0.05, cP<0.01 vs control cells.
Discussion
Curcumin interacts with a variety of proteins and modifies their expression and activity; thus, it is a good candidate preventive agent and/or therapeutic agent for cancer. Moreover, curcumin reduces the toxicity induced by anti-cancer agents, sensitizes chemo-resistant cancer cells and demonstrates synergic effects with different chemotherapeutic agents, such as doxorubicin, paclitaxel, vincristine, cisplatin, gemcitabine, oxaliplatin, etoposide, thalidomide, etc22. It was previously reported that curcumin suppresses proliferation and induces apoptosis in a variety of tumor cells; however, the molecular mechanisms underlying these effects are still unclear.
The data presented in this article show that curcumin treatment caused a significant dose- and time-dependent decrease in NTera-2 cell proliferation compared with untreated controls. The IC50 values for curcumin in NTera-2 cells at 8, 16, 24, and 32 h were 12.46, 16.97, 10.70, and 8.87 μmol/L, respectively. Moreover, curcumin treatment suppressed colony formation in all of the testicular cancer cell lines tested, suggesting that curcumin's effect on the tumor cells is irreversible. We demonstrated that curcumin induced apoptosis in the human malignant testicular germ cell line, NTera-2, by regulating AP-2γ and its downstream survival signaling pathways. Curcumin induced apoptosis in a dose-dependent manner. After NTera-2 cells were exposed to the indicated doses of curcumin for 24 h, nuclear morphological changes typical of apoptosis were observed with Hoechst 33258 staining, and there was an increase in the number of TUNEL-positive cells. To determine whether curcumin-induced cell death was due to apoptosis or necrosis, annexin V/PI double labeling assays were performed. The treatment of NTera-2 cells with various concentrations of curcumin (5, 10, and 15 μmol/L) resulted in apoptosis rates of 7.8%±0.58%, 13.0%±1.06%, and 18%±1.78%, respectively, which were significantly higher than that of the control group (4.6%±0.31%) (Figure 2C). Thus, the induction of NTera-2 cell apoptosis in response to curcumin is an important mechanism for its preventative and anti-tumor activity in testicular cancer.
Apoptosis occurs via a mitochondrial-dependent intrinsic pathway or a death receptor-mediated extrinsic pathway23,24,25. The intrinsic pathway of apoptosis is regulated by members of the Bcl-2 family, which is composed of proapoptotic members (ie, Bax, and Bad) and anti-apoptotic members (ie, Bcl-2 and Bcl-XL). In contrast, so-called “extrinsic” pathway signals, such as those mediated by death receptors of the TNF receptor superfamily, directly activate the caspase cascade. For example, interaction of Fas with its ligand (FasL) triggers the formation of a death-inducing signaling complex (DISC) that includes the critical adaptor molecule FADD, which recruits procaspase-8. It was reported that the activation of caspases contributed to curcumin-induced apoptosis in breast tumors. Similarly, we found that caspase 9, 8, and 3 cleavage play a role in curcumin-induced apoptosis in NTera-2 cells. Moreover, the expression of Bcl-2 and FasL protein were simultaneously decreased, and Bax levels were up-regulated following curcumin treatment in NTera-2 cells. These data indicate that the intrinsic and extrinsic pathways both play an important role in curcumin-induced apoptosis in NTera-2 cells.
Advances in the understanding of the molecular mechanisms of apoptosis have identified the apoptotic pathway as a promising target to increase the effectiveness of cancer treatment. A recent study revealed a dual role for AP-2γ in different mammary tumorigenic stages: inhibition of tumor initiation and promotion of proliferation. These findings provide evidence that AP-2γ participates in complex biological processes, including cell cycle progression, apoptosis, and tumor formation. Increased AP-2γ expression is associated with poor prognosis in patients with germ cell tumors, and therefore, it represents a target for intervention. Accordingly, we hypothesized that curcumin might exert its anticancer effects via modulation of AP-2γ expression. Experimental data confirmed that, in vitro, AP-2γ activity is linked to its expression level, which suggests that the regulation of AP-2γ expression is critical in tumorigenesis. Thus, drugs that can downregulate AP-2γ could have potential therapeutic value. The results presented here demonstrate that curcumin directly inhibits AP-2γ expression in a dose-dependent manner in NTera-2 cells. Therefore, because curcumin can block the expression of AP-2γ, it may represent a useful treatment for AP-2γ overexpressing testicular cancer. Western blotting assays revealed that siRNA-mediated knockdown of AP-2γ induced the cleavage of apical pro-caspase 3, pro-caspase 8, pro-caspase 9, and the subsequent proteolytic cleavage of PARP in NTera-2 cells. This experiment indicated that down-regulation of AP-2γ alone was sufficient to increase apoptosis in NTera-2 cells. Moreover, the effects of-AP-2γ knockdown were consistent with those of curcumin treatment in NTera-2 cells. Therefore, we conclude that curcumin regulates NTera-2 cell apoptosis by targeting AP-2γ.
Curcumin restrains cancer cell progression via telomerase activity suppression. However, the exact mechanism is still unknown. The proteasomal degradation pathway is essential for many cellular processes, including the cell cycle, the regulation of gene expression, and the oxidative stress response. Curcumin has been shown to inhibit activities of the proteasome in certain cancer cells26; however, Fossey et al found that no evidence for this activity after treating the OSA cell lines with curcumin at the doses and time points examined27. Moreover, previous studies reported that the decreased expression of Sp1 in A549 cells and cyclin E in MCF-7 cells by curcumin were reversed by proteasome inhibitors, suggesting the role of ubiquitin-dependent proteasomal pathway28,29. In the present study, we explored the mechanism by which curcumin reduces AP-2γ protein levels in NTera-2 cells. As shown in Figure 5, AP-2γ was much more rapidly degraded in NTera-2 cells after treatment with curcumin. However, treatment with the proteasome inhibitor, MG132, efficiently rescued the protein level of AP-2γ, indicating that AP-2γ is degraded in a ubiquitin-proteasome-dependent manner. In addition, MTT assays revealed that the proteasome inhibitor, MG132, reversed the curcumin-mediated inhibition of proliferation in the same condition. These results suggest that curcumin can increase the proteasome-dependent degradation of AP-2γ. However, AP-2γ is not the only pathway active in testicular cancer, as many other signal transduction pathways are activated, and these pathways will be the focus of our future research.
AP-2 is a strong activator of the ErbB2 gene, and it is functionally activated in ErbB2-overexpressing breast cancer cell lines30. The molecular mechanisms leading to ErbB2 overexpression are increased gene amplification and transcription. Various experimental approaches have shown that the loss of functional ErbB2 in tumor cells causes growth inhibition and induces apoptosis. Increased ErbB2 expression is correlated with the presence of one or more AP-2 proteins, especially AP-2a and AP-2γ. The aims of this study were to investigate the effects of curcumin on AP-2γ and to elucidate its underlying mechanisms of action. Our examinations revealed that treatment of NTera-2 cells with curcumin inhibited the expression of AP-2γ and decreased ErbB2 protein expression. In conclusion, blocking the expression of ErbB2 with curcumin may partly inhibit the expression of AP-2γ. The role of other transcription factors will be studied in the future. The transforming effects of ErbB2 receptors are mediated through the well-characterized MAPK-ERK and PI3K-AKT pathways. Recent data show that the MAPK pathway plays critical roles in cell survival and death in many physiological and pathological settings31,32. In the present study, changes in the phosphorylation of ERK and AKT were examined using phospho-specific antibodies following curcumin treatment in NTera-2 cells. Further studies using specific inhibitors of ERK and AKT are required to determine whether ERK and AKT activity are essential for the anticancer activity of curcumin. Other AP-2γ-controlled genes are also likely to be involved in this pathway, and they will need to be further explored in future studies. Our data suggest that curcumin inhibits cell growth and induces apoptosis, at least in part, via the inactivation of AP-2γ, which is known to play a significant role in cancer cell proliferation.
In summary, we established an in vitro model to evaluate the efficacy of curcumin as a therapy for testicular cancer and investigated its mechanisms of action. Our preliminary observations indicate that curcumin arrests malignant testicular germ cell proliferation and induces cell apoptosis, at least in part, via the caspase-dependent apoptotic pathway, degradation of AP-2γ and the down-regulation of ERK/AKT signaling. Thus, targeting AP-2γ may be an effective strategy to control tissue destruction in testicular cancer patients.
Author contribution
Chang ZHOU and Jian ZHANG conceived and designed the experiments. Xiao-meng ZHAO, Cheng WANG, Xiao-ting ZHANG, and Xiao-feng LI performed the experiments. Chang ZHOU and Xiao-meng ZHAO analyzed the data. Xiao-feng DING, Xi-zhi LIU, and Shuang-lin XIANG contributed reagents/materials/analysis tools. Chang ZHOU and Xiao-meng ZHAO wrote the paper. All authors discussed the results and commented on the manuscript.
References
Iczkowski KA, Butler SL . New immunohistochemical markers in testicular tumors. Anal Quant Cytol Histol 2006; 28: 181–7.
Holzik MF, Sijmons RH, Josette EHM, Hoekstra-Weebers, Sleijfer DT, Hoekstra HJ . Clinical and genetic aspects of testicular germ cell tumours. Hereditary Cancer Clin Practice 2008; 6: 3–14.
Aggarwal BB, Kumar A, Aggarwal MS, Shishodia S . Curcumin derived from turmeric (Curcuma longa): a spice for all seasons. Phytopharm Cancer Chemopreven 2005; 349–87.
Anand P, Sundaram C, Jhurani S, Kunnumakkara AB, Aggarwal BB . Curcumin and cancer: an “old-age” disease with an “age-old"”solution. Cancer Lett 2008; 267: 133–64.
Kunnumakkara AB, Anand P, Aggarwal BB . Curcumin inhibits proliferation, invasion, angiogenesis and metastasis of different cancers through interaction with multiple cell signaling proteins. Cancer Lett 2008; 269: 199–225.
Hartojo W, Silvers AL, Thomas DG, Seder CW, Lin L, Rao H, et al. Curcumin promotes apoptosis, increases chemosensitivity, and inhibits nuclear factor kappaB in esophageal adenocarcinoma. Transl Oncol 2010; 3: 99–108.
Subramaniam D, Ponnurangam S, Ramamoorthy P, Standing D, Battafarano RJ, Anant S, et al. Curcumin induces cell death in esophageal cancer cells through modulating Notch signaling. PLoS One 2012; 7: e30590.
Ravindran J, Prasad S, Aggarwal BB . Curcumin and cancer cells: how many ways can curry kill tumor cells selectively? AAPS J 2009; 11: 495510.
Shakibaei M, John T, Schulze-Tanzil G, Lehmann I, Mobasheri A . Suppression of NF-kappaB activation by curcumin leads to inhibition of expression of cyclo-oxygenase-2 and matrix metalloproteinase-9 in human articular chondrocytes: Implications for the treatment of osteoarthritis. Biochem Pharmacol 2007; 3: 1434–45.
Csaki C, Mobasheri A, Shakibaei M . Synergistic chondroprotective effects of curcumin and resveratrol in human articular chondrocytes: inhibition of IL-1beta-induced NF-kappaB-mediated inflammation and apoptosis. Arthritis Res Ther 2009; 11: R165.
Werling U, Schorle H . Transcription factor gene AP-2γ essential for early murine development. Mol Cell Biol 2002; 22: 3149–56.
Hoei-Hansen CE, Nielsen JE, Almstrup K, Sonne SB, Graem N, Skakkebaek NE, et al. Transcription factor AP-2gamma is a developmentally regulated marker of testicular carcinoma in situ and germ cell tumors. Clin Cancer Res 2004; 10: 8521–30.
Anttila MA, Kellokoski JK, Moisio KI, Mitchell PJ, Saarikoski S, Syrjänen K, et al. Expression of transcription factor AP-2alpha predicts survival in epithelial ovarian cancer. Br J Cancer 2000; 82: 1974–83.
Provenzano MJ, Yu L, Hitchler MJ, Fitzgerald MP, Robinson RA, Wayne S, et al. AP-2 participates in the transcriptional control of the amyloid precursor protein (APP) gene in oral squamous cell carcinoma. Exp Mol Pathol 2007; 83: 277–82.
Ailan H, Xiangwen X, Daolong R, Lu G, Xiaofeng D, Xi Q, et al. Identification of target genes of transcription factor activator protein 2 gamma in breast cancer cells. BMC Cancer 2009; 9: 279
Jäger R, Werling U, Rimpf S, Jacob A, Schorle H . Transcription factor AP-2gamma stimulates proliferation and apoptosis and impairs differentiation in a transgenic model. Mol Cancer Res 2003; 1: 921–9.
Li H, Goswami PC, Domann FE . AP-2γ Induces p21 expression, arrests cell cycle, and inhibits the tumor growth of human carcinoma cells. Neoplasia 2006; 8: 568–77.
Hoei-Hansen CE, Carlsen E, Jorgensen N, Leffers H, Skakkebaek NE, Rajpert-De Meyts E . Towards a non-invasive method for early detection of testicular neoplasia in semen samples by identification of fetal germ cell-specific markers. Hum Reprod 2007; 22: 167–73.
Xiang S, Sun Z, He Q, Yan F, Wang Y, Zhang J . Aspirin inhibits ErbB2 to induce apoptosis in cervical cancer cells. Med Oncol 2010; 27: 379–87.
Droga-Mazovec G, Bojic L, Petelin A, Ivanova S, Romih R, Repnik U, et al. Cysteine cathepsins trigger caspase-dependent cell death through cleavage of bid and antiapoptotic Bcl-2 homologues. J Biol Chem 2008; 283: 19140–50.
Wagner EF, Nebreda AR . Signal integration by JNK and p38 MAPK pathways in cancer development. Nat Rev Cancer 2009; 9: 537–49.
Nautiyal J, Banerjee S, Kanwar SS, Yu Y, Patel BB, Sarkar FH, et al. Curcumin enhances dasatinib-induced inhibition of growth and transformation of colon cancer cells. Int J Cancer 2011; 128: 951–61.
Heath-Engel HM, Chang NC, Shore GC . The endoplasmic reticulum in apoptosis and autophagy: role of the BCL-2 protein family. Oncogene 2008; 27: 6419–33.
Yun SI, Yoon HY, Chung YS . Glycogen synthase kinase-3beta regulates etoposide-induced apoptosis via Bcl-2 mediated caspase-3 activation in C3H10T1/2 cells. Apoptosis 2009; 14: 771–7.
Droga-Mazovec G, Bojic L, Petelin A, Ivanova S, Romih R, Repnik U, et al. Cysteine cathepsins trigger caspase-dependent cell death through cleavage of bid and antiapoptotic Bcl-2 homologues. J Biol Chem 2008; 283: 19140–50.
Milacic V, Banerjee S, Landis-Piwowar KR, Sarkar FH, Majumdar AP, Dou QP . Curcumin inhibits the proteasome activity in human colon cancer cells in vitro and in vivo. Cancer Res 2008; 68: 7283–92.
Fossey SL, Bear MD, Lin J, Li C, Schwartz EB, Li PK, et al. The novel curcumin analog FLLL32 decreases STAT3 DNA binding activity and expression, and induces apoptosis in osteosarcoma cell lines. BMC Cancer 2011; 11: 112.
Hsin IL, Sheu GT, Chen HH, Chiu LY, Wang HD, Chan HW, et al. N-acetyl cysteine mitigates curcumin-mediated telomerase inhibition through rescuing of Sp1 reduction in A549 cells. Mutat Res 2010; 688: 72–7.
Aggarwal BB, Banerjee S, Bharadwaj U, Sung B, Shishodia S, Sethi G . Curcumin induces the degradation of cyclin E expression through ubiquitin-dependent pathway and up-regulates cyclin-dependent kinase inhibitors p21 and p27 in multiple human tumor cell lines. Biochem Pharmacol 2007; 73: 1024–32.
Woodfield GW, Chen Y, Bair TB, Domann FE, Weigel RJ . Identification of primary gene targets of TFAP2C in hormone responsive breast carcinoma cells. Genes Chromosomes Cancer 2010; 49: 948–62.
Tari AM, Lopez-Berestein G . Serum predominantly activates MAPK and akt kinases in EGFR- and ErbB2-over-expressing cells, respectively. Int J Cancer 2000; 86: 295–7.
Yarden Y, Sliwkowski MX . Untangling the ErbB signalling network. Nat Rev Mol Cell Biol 2001; 2: 127–37.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (Grant Nos 81071696 and 31071150), Project of Hunan Provincial Masters Innovation Fund (CX2012B227) and the Project of Chang Sha Science and Technology Plan (K1109006-31, K1207014-31).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article
Cite this article
Zhou, C., Zhao, Xm., Li, Xf. et al. Curcumin inhibits AP-2γ-induced apoptosis in the human malignant testicular germ cells in vitro. Acta Pharmacol Sin 34, 1192–1200 (2013). https://doi.org/10.1038/aps.2013.38
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/aps.2013.38
Keywords
This article is cited by
-
Exploring the role of natural bioactive molecules in genitourinary cancers: how far has research progressed?
Natural Products and Bioprospecting (2023)
-
Tetramethylpyrazine Protects Against Oxygen-Glucose Deprivation-Induced Brain Microvascular Endothelial Cells Injury via Rho/Rho-kinase Signaling Pathway
Cellular and Molecular Neurobiology (2017)
