
Abstract
Clinical and biochemical findings in 5 unrelated patients with osteogenesis imperfecta (OI) with a serine for glycine substitution in the α2(I) collagen chain are presented. The data are compared to other serine substitutions in collagen type I. Findings show that the phenotypic severity of serine for glycine substitutions in the α2(I) collagen chain is region dependent similar to the observations for the α1(I) collagen chain, and that so-called ‘lethal’ and ‘non-lethal’ domains in the α1 and α2 collagen chains do not necessarily correspond.
Similar content being viewed by others

Introduction
Osteogenesis imperfecta (OI) is a heterogeneous connective tissue disorder characterized by brittle bones. Four clinical subtypes are recognized, including a mild (type I), a lethal (type II), a severely deforming (type III) and a moderately severe (type IV) OI subtype [1]. A wide range of underlying mutations in COL1A1 and COL1A2 — the genes encoding both type I collagen α-chains — are known. These comprise glycine substitutions, splicing mutations causing exon skipping and, more rarely, large gene deletions [2].
The phenotypic outcome of glycine substitutions in type I collagen is influenced by the position and the nature of the substituting residue [2, 3]. It has been shown previously that glycine substitutions in the α1(I) collagen chain generally show a linear gradient of phenotypic severity from the carboxyl- to the amino-terminal end of the molecule. However, as an increasing number of glycine substitutions in type I collagen is identified, more and more exceptions to this model are encountered. In this paper, we report 5 unrelated OI probands with glycine to serine substitutions in the α2(I) collagen chain and compare the phenotypic outcome with that of other serine substitutions reported to the Osteogenesis Imperfecta Mutation Consortium.
Materials and Methods
Clinical Summaries
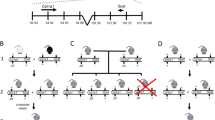
Proband AU (fig. 1A) is a male fetus, second child of healthy, non-consanguineous parents. The pregnancy was terminated at 21 weeks of gestation after detection of multiple fractures on ultrasound. One elder sib is healthy. Clinical examination revealed a very small baby with a vertex to sacrum length of 16.6 cm and a weight of 390 g. There was shortening of the upper and lower limbs, marked bowing and deformity of the lower legs and abduction of the thighs.

A Babygram of patient AU with the α2(I)-G859S substitution showing poor ossification of the calvarium, thin ribs, mild vertebral platyspondyly, severe shortening of the femora and angulation of tibiae and fibulae. B X-ray of patient PT with the α2(I)-G811S substitution, showing osteopenia, mild bowing of the femora and some widening of the metaphyses. C X-ray of patient WN with the α2(I)-G682S substitution showing osteopenia and bowing of the femora.
Radiological features showed a thin, poorly ossified calvarium, thin ribs with no visible fractures, and some platyspondyly of the vertebrae. The femora were short and irregular, the tibiae broad and angulated, the fibulae very thin and angulated. Several fractures of the humeri and ulnae were seen. The clinical and radiological features were those of OI type IIB/III.
Proband PT (fig. 1B) is the first child of healthy, non-consanguineous parents, born at 40 weeks gestation after a breech delivery. Birth length was below the 3rd percentile. At birth, she showed several fractures and presented with a large head, triangular face, small nose, moderate hypotonia, enlarged fontanelles and white sclerae. Radiological features included relatively adequate ossification of the calvarium and multiple wormian bones, small, thin ribs and some platyspondyly of the cervical and thoracic vertebrae. The long bones were short but adequately modelled; there was some bowing and metaphyseal flaring of the humeri and tibiae. The femora were short and bowed, the fibulae were very thin. The clinical and radiological data fitted with OI type IV.
The proband’s mother and one maternal uncle have multiple tooth caries and dentinogenesis imperfecta. One cousin has short stature and suffered several fractures. Two cousins of the proband’s mother were diagnosed with Camurati-Engelman disease.
Proband WN (fig. 1C) is the second child of non-consanguineous healthy parents, born after an uneventful pregnancy. One older brother is healthy. A second-degree cousin is deaf-mute. The patient suffered her first fracture (right femur) at age 2½ years, the second fracture (left tibia) at age 8. Between age 8 and 10 years, she suffered 4 additional fractures of the right femur. She could not walk for 3 years and is now using crutches. Radiologic evaluation at age 13 showed marked osteopenia with thin cortices and varus deformity and bowing of the femora. She suffered 6 more fractures between age 11 and 16 years. Now, at age 26, she has developed a severe thoracic and lumbar scoliosis. Her stature is 152 cm, her sclerae are white and hearing is normal. Her phenotype is compatible with OI type IV.
Proband MJ (fig. 2A) was first seen by us at age 40. She was born at term after an uneventful pregnancy. Reportedly, she suffered her first fracture at 3½ months. She had severe difficulties walking since early childhood, moved with crutches at age 10 and became wheelchair-bound at age 14. She suffered more than 40 fractures, mostly of the lower extremities. The fractures were treated with casts and always involved long periods of immobilisation. By age 38, her hearing had decreased.

Clinical and X-rays of patients MJ and KJ with a α2(I)-G238S substitution. A Patient MJ at age 45, showing severe osteopenia and deformation of pelvis and the long bones of the lower extremities. B Patient KJ at age 4, already showing distinct bowing of the femora and tibiae.
Clinical evaluation at age 40 showed that she was severely dwarfed with a stature below 1.40 m and presented with severe shortening and bowing of her lower extremities. In addition, she had a large head with triangular face, white sclerae, opalescent teeth, joint laxity, severe thoracic hyperkyphosis and lumbar hyperlordosis and flexion contractures of the hips. In contrast, the humeri appeared relatively well modelled and less short. She now suffers from digestive problems due to a huge herniation of the bowel into the thoracic cage. Her phenotype is compatible with OI type III.
Proband KJ (fig. 2B) is the first child of healthy, non-consanguineous parents. The diagnosis of OI was made on ultrasound in the 7th month of pregnancy. Born at term, he presented two fractures and a birth length of 48 cm. He suffered at least 6 more fractures in the first years of life, had delayed motor development and walked independently by age 3. Examination at that time showed a stature below the 3rd percentile, a large turricephalic skull, enlarged fontanelles, slightly blue sclerae, low-set ears, dentinogenesis imperfecta, short extremities and mild joint hyperlaxity. Radiological examination disclosed multiple wormian bones, several old rib fractures, very short humeri and mild shortening and anterior bowing of the femora and tibiae. Now, at age 4, he is walking independently without physical impairment. His phenotype is OI type IV.
Biochemical Studies
Skin biopsies were obtained from the 5 OI probands (AU, PT, WN, KJ, MJ), and from the parents of patients WN and AU. Fibroblast cultures were established under standard conditions. After labelling with 14C-proline, the procollagen and collagen proteins were isolated from medium and cell layer and examined by SDS electrophoresis as described earlier [4]. Thermal stability measurements were performed as described by Bruckner and Prockop [5]. Collagen secreted into the medium was subjected to peptide mapping by in situ cyanogen bromide digestion followed by electrophoresis in the second dimension [6]. Fibroblast cultures labelled in the presence of 0.3 mM αα′-dipyridyl were harvested the same way and evaluated by SDS-PAGE.
Molecular Investigations
Total RNA was isolated from cultured fibroblasts, and cDNA was obtained, using M-MLV reversed transcriptase (Life Technologies) according to the manufacturer’s instructions. PCR was performed using primers for the COL1A2 coding region and the amplimers were analyzed by single strand conformation polymorphism analysis (SSCP) as described [7] or by heteroduplex analysis using conformation-sensitive gel electrophoresis (CSGE) as described [8].
Fragments showing an abnormal migration pattern were cloned using pCR-Script cloning kit (Stratagene) and sequenced by the dideoxy chain termination method [9] (numbering starts from the first Gly residue in the α2(I) collagen chain as described in the EMBL sequence Z74616).
The presence of the mutation was investigated by restriction enzyme digestion on cDNA and, where possible, on genomic DNA from the patients and the parents. Genomic DNA was isolated from peripheral blood leukocytes using the Qiagen-Blood miniprep kit (Qiagen Inc. Chatworth, Calif., USA).
Results
Biochemical Collagen Analysis
SDS electrophoresis of pepsin-digested collagen from the medium showed slightly altered collagen profiles for each of the probands, compared to the control samples (fig. 3A). In patients AU, WN, PT and KJ, broadening of the bands representing the α1-and/or α2-chains of type I collagen was seen, suggesting that in addition to normal α-chains a population of slowly migrating overmodified αM-chains was synthesized. However, the degree of overmodification was much more pronounced in AU than in the other OI probands. In patients WN and MJ, the intensity of the α2(I) band was decreased and in patient WN, a mild delay in electrophoretic mobility of the α2(I) chains was visible. Analysis of the cell layer collagens (fig. 3B) revealed patterns similar to the medium collagens, except in patient MJ, in whom a mild broadening of the α2(I) chains was seen, compatible with retention of mutant collagen chains in the cells.

Analysis by SDS-PAGE of pepsin-derived collagens secreted by cultured fibroblasts. Lane 1: control samples; lane 2: patient MJ; lane 3: patient KJ; lane 4: patient WN; lane 5: patient PT, and lane 6: patient AU. α1(I) and α2(I): normal collagen type I α-chains; α1(I)m and α2(I)m: mutant, more slowly migrating collagen type I α-chains. A Electrophoresis of the collagen secreted into the medium showed broadening of the bands representing the α1(I) and α2(I) chains in all patients, except in patient MJ. The band representing the α2(I) chains was decreased in intensity in patients MJ (lane 2) and WN (lane 4), and showed delayed migration in patient WN. B Cell-layer collagens derived from the 5 OI patients showed varying broadening of the bands representing the collagen type I α-chains. In patient MJ, broadening of the collagen α-chains is seen in contrast to the normal findings in the medium, suggesting retention of the mutant collagen type I chains in the cell layer.
Analysis of the procollagens showed a variable delay in processing of procollagen type I in the 5 probands as compared to controls (data not shown).
SDS-PAGE of collagens extracted from fibroblast cultures labelled in the presence of αα′-dipyridyl showed that the slow-migrating type I collagen α-chains disappeared (data not shown), indicating that enzymatic overmodification was indeed the cause of the delayed mobility. The collagen type I melting profiles showed a slight decrease of 1–2°C in denaturing temperature in all OI probands (table 1).
In situ two-dimensional peptide mapping of the type I collagen chains showed that enzymatic overmodification started in the α1(I) CB6 peptide for patient AU, in the α1(I) CB7 peptide for patients PT and WN and in the α1(I) CB8 peptide for patient KJ, suggesting that either the mutation was localized in one of those α1(I) CB peptides or to the corresponding region of the α2(I) chain (data not shown).
Molecular Analyses
Using primers covering the complete COL1A1 and COL1A2 α-helix coding region, RT-PCR was performed on mRNA derived from the patients’ fibroblasts. Migration shifts were detected in the COL1A2 gene by SSCP in the patients MJ, WN and AU and by heteroduplex analysis in patients KJ and PT. Cloning and sequencing of the relevant COL1A2 fragments resulted in the identification of 4 different G to A transitions, which changed the codon for glycine to that for serine (table 1): a G238S in patients MJ and KJ, a G682S in patient WN, a G811S in patient PT and a G859S in patient AU. The presence of the mutation was confirmed by restriction analysis on cDNA in the 5 OI patients, and on genomic DNA isolated from leukocytes in patients MJ, KJ and WN. No mutation was found in the parents of patients MJ, KJ and WN. Genomic DNA from all family members of patient PT were analyzed by restriction digestion, but the α2-G811S substitution was not present.
Discussion
Substitutions of serine for glycine in collagen type I are the most common single base substitutions detected in OI. So far, the number of serine substitutions identified in the α1(I) chain by far exceeds that of serine substitutions in the α2(I) collagen chain, as shown by the data of the Osteogenesis Imperfecta Mutation Consortium (OIMC) (fig. 4).

Diagram summarizing serine for glycine substitutions in the α1(I) and α2(I) collagen chain, as reported to the OIMC (September 1996).
The 5 OI probands described here all harbor substitutions of serine for glycine in the α2(I) collagen chain, resulting from a heterogeneous G → A transition at a CpG nucleotide. Pruchno et al. [10] have suggested that CpG dinucleotides within the coding region of the type I collagen genes represent ‘hot spots’ for mutation. This is supported by the fact that all recurrent mutations described so far in OI involve CpG dinucleotides.
In the 5 patients described here, the OI phenotype most probably results from a new mutation since in all of them the family history is negative. In particular, in patients MJ, KJ and WN, no evidence for mosaicism was found as a possible cause for the differences in phenotypic severity in these patients.
The biochemical profiles are in agreement with the degree of overmodification as expected from the position of the substitution of the serine for glycine along the α2(I) collagen chain. In 2 patients, MJ and WN, a reduced amount of α2(I) collagen chains was observed. In the first, MJ, this can be explained by intracellular retention of the mutant α2(I) collagen chains (fig. 3B). In patient WN, there is no evidence for intracellular accumulation, but rapid degradation of mutant α2(I) chains may explain the reduction of α2(I) collagen chains in the medium.
The serine substitutions span a major part of the α2(I) collagen helix. The substitution closest to the C terminus, G859S, causes the most severe OI phenotype, whereas G811S and G682S produce a moderately severe phenotype and the N-terminal G238S substitution is associated with a moderately (patient KJ) and a severely deforming (patient MJ) OI phenotype. Three of the serine substitutions have been reported earlier in OI patients. The G859S substitution was previously detected in a patient with the severe OI type III phenotype, as seen in patient AU [11]. The G811S substitution causing moderately severe OI in patient PT was previously documented in a patient with lethal OI [Byers et al., pers. commun.]. The G238S substitution (patients KJ and MJ) was reported respectively in patients with OI type III [12], OI type I and OI type I/IV [Byers et al., pers. commun.]. This mutation may represent a ‘hot spot’, as it has also been detected in several other OI patients [Byers et al., pers. commun.]. Differences in phenotypic severity resulting from the same glycine substitution have been detected not only in unrelated OI patients [13, 14] but also in patients from the same family [15]. An explanation for these observations is not found yet, but variation in genetic background as well as differences in levels of gene expression may be involved. Dyne et al. [16] described deficient expression of decorin in a case of severe/lethal OI presenting a α1(I)-G415S substitution. The clinical outcome in this particular patient was much more severe compared to 2 other OI patients with the same substitution but with normal decorin expression levels. Sakai et al. [17] discussed the role of as yet unidentified ‘modifiers’ in the molecular, intra- and extracellular pathways as factors influencing phenotypic variability. In addition to this, differences in therapeutic management have most probably also contributed to some extent to the phenotypic differences between patients KJ and MJ. Indeed, the long periods of immobilisation and lack of an adequate physical rehabilitation programme most certainly worsened the physical condition of patient MJ as compared to patient KJ, who has received early intensive orthopaedic care.
The mutations reported here add to the data suggesting that although a gradient of clinical severity from the carboxyl- to amino-terminal end is generally observed for collagen type I mutations, there are exceptions to this model, in particular for glycine to serine substitutions irrespective of the type of chain involved.
Three substitutions of serine for glycine have been identified as well in the α1(I) as in the α2(I) collagen chain, i.e. the G247S, G631S and G661S substitutions [7, 18–21; Byers et al., pers. commun.]. The α1(I) G247S and G661S substitutions cause distinctly more severe phenotypes than their homologues in the α2-chain, which is explained by the fact that the type I collagen molecule contains two α1-chains and one α2-chain. However, this is not the case for the G631S substitution which causes a lethal phenotype in all instances [19; Byers et al., pers. commun.]. This illustrates that besides the chain effect, other domain-related factors determine the phenotypic outcome of a glycine substitution in type I collagen. There are ‘lethal’ regions which are crucial for the structural and functional integrity of the molecule and ‘non-lethal’ regions, in which sequence variations have a less dramatic effect on the molecular stability function. Apparently, these regions do not necessarily correspond for both collagen type I α-chains, suggesting that these regions are chain specific, rather than molecule specific. A possible explanation for this phenomenon is the specific chain bonding of yet unidentified molecules which play a role in the intra- and extracellular pathways and hence contribute to correct fibril formation and function.
In conclusion, the phenotypic outcome of collagen type I mutations, as illustrated here for glycine to serine substitutions, is influenced by many different factors. Besides the position and nature of the mutation, other effects such as epigenetic factors and environmental influences determine the phenotypic outcome and make it difficult to predict the OI phenotype from the genotype.
References
Sillence DO, Senn A, Danks DM: Genetic heterogeneity in osteogenesis imperfecta. J Med Genet 1979;16:101–116.
Byers PH: Osteogenesis imperfecta; in Royce PM, Steinmann B (eds): Connective tissue and its heritable disorders: Molecular, genetic and medical aspects. New York, Wiley-Liss, 1993, pp 317–350.
Kuivaniemi H, Tromp G, Prockop DJ: Mutations in collagen genes: Causes of rare and some common diseases in humans. FASEB J 1991;5: 2052–2060.
Nuytinck L, Narcisi P, Nicholls A, Renard JP, Pope FM, De Paepe A: Detection and characterisation of an overmodified type III collagen by analysis of non-cutaneous connective tissues in a patient with Ehlers-Danlos syndrome IV. J Med Genet 1992;29:375–380.
Bruckner P, Prockop DJ: Proteolytic enzymes as probes for the triple-helical conformation of procollagen. Anal Biochem 1981; 110:360–368.
Barsh GS, Peterson KE, Byers PJ: Peptide mapping of collagen chains using CNBr cleavage of proteins within Polyacrylamide gels. Collagen Rel Res 1981;1:543–548.
Mackay K, Byers PH, Dalgleish R: An RTPCR-SSCP strategy for detection of mutations in the gene encoding the α1 chain of type I collagen: Application to four patients with osteogenesis imperfecta. Hum Mol Genet 1993;2: 1155–1160.
Ganguly A, Rock M, Prockop DJ: Conformation-sensitive gel electrophoresis for rapid detection of single-base differences in double-stranded PCR products and DNA fragments: Evidence for solvent-induced bends in DNA heteroduplexes. Proc Natl Acad Sci USA 1993; 90:10325–10329.
Sanger R, Nicklen FS, Coulson AR: DNA sequencing with chain-termination inhibitors. Proc Natl Acad Sci USA 1979;74:5463–5467.
Pruchno CJ, Cohn DH, Wallis GA, Willing MC, Starman BJ, Zhang X, Byers PH: Osteogenesis imperfecta due to recurrent point mutations at CpG dinucleotides in the COL1A1 gene of type I collagen. Hum Genet 1991;87: 33–40.
Rose NJ, Mackay K, Byers PH, Dalgleish R: A Gly859Ser substitution in the triple helical domain of the α2 chain of type I collagen resulting in osteogenesis imperfecta type III in two unrelated individuals. Hum Mut 1994;3:391–394.
Rose NJ, Mackay K, Byers PH, Dalgleish R: A Gly238Ser substitution in the α2 chain of type I collagen results in osteogenesis imperfecta type III. Hum Genet 1995;95:215–218.
Cole WG, Dalgleish R: Perinatal lethal osteogenesis imperfecta. J Med Genet 1995;32:284–289.
Mottes M, Gomez Lira MM, Valli M, Scarano G, Lonardo F, Forlino A, Cetta G, Pignatti PF: Paternal mosaicism for a COL1A1 dominant mutation (al Ser-415) causes recurrent osteogenesis imperfecta. Hum Mut 1993;2:196–204.
Superti-Furga A, Pistone F, Romano C, Steinmann B: Clinical variability of osteogenesis imperfecta linked to COL1A2 and associated with a structural defect in the type I collagen molecule. J Med Genet 1989;26:358–362.
Dyne KM, Valli M, Forlino A, Mottes M, Kresse H, Cetta G: Deficient expression of the small proteoglycan decorin in a case of severe/lethal osteogenesis imperfecta. Am J Med Genet 1996;63:161–166.
Sakai LY, Burgeson RE, Olsen BR, Rowe DW, Gordon SL: Current knowledge and research directions in heritable disorders of connective tissue. Matrix Biol 1996; 15:211–229.
Zhuang J, Tromp G, Kuivaniemi H, Castells S, Bugge M, Prockop DJ: Direct sequencing of PCR products derived from cDNAs for the proα1 and proα2 chains of type I procollagen as a screening method to detect mutation in patients with osteogenesis imperfecta. Hum Mut 1996;7:89–99.
Westerhausen A, Kishi J, Prockop DJ: Mutations that substitute serine for glycine α1-598 and glycine α1-631 in type I procollagen. J Biol Chem 1990;265:13995–14000.
Spotila LD, Constantinou CD, Sereda L, Ganguly A, Riggs BL, Prockop DJ: Mutation in a gene for type I procollagen (COL1A2) in a woman with postmenopausal osteoporosis: Evidence for phenotypic and genotypic overlap with mild osteogenesis imperfecta. Proc Natl Acad Sci USA 1991;88:5423–5427.
Nuytinck L, Dalgleish R, Spotila L, Renard JP, Van Regemorter N, De Paepe A: Substitution of glycine-661 by serine in the α1(I) and α2(I) chains of type I collagen results in different clinical and biochemical phenotypes. Hum Genet 1996;97:324–329.
Acknowledgements
We are grateful to J.P. Renard and M. Van Thielen for excellent technical assistance. We are endebted to Dr. P. Byers and Dr. A. Lund for permission to report results communicated to the Osteogenesis Imperfecta Mutation Consortium but not yet published. This work was supported by the Fund of Scientific Research, Flanders, Belgium.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Nuytinck, L., Wettinck, K., Freund, M. et al. Osteogenesis imperfecta Phenotypes Resulting from Serine for Glycine Substitutions in the α2(I) Collagen Chain. Eur J Hum Genet 5, 161–167 (1997). https://doi.org/10.1007/BF03405894
Received:
Revised:
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF03405894
